BlaR1 Fragmentation and Proteasomal Turnover: The Key Mechanism for β-Lactam Antibiotic Signal Recovery and Resistance
This article provides a comprehensive analysis of BlaR1, the signal-transducing membrane sensor for β-lactam antibiotics, focusing on its critical fragmentation and proteasome-mediated degradation as the central mechanism for signal recovery.
BlaR1 Fragmentation and Proteasomal Turnover: The Key Mechanism for β-Lactam Antibiotic Signal Recovery and Resistance
Abstract
This article provides a comprehensive analysis of BlaR1, the signal-transducing membrane sensor for β-lactam antibiotics, focusing on its critical fragmentation and proteasome-mediated degradation as the central mechanism for signal recovery. We explore the foundational biology of BlaR1 activation, detail the methodological approaches used to study its regulated turnover, address common experimental challenges, and validate findings through comparative analysis with other resistance mechanisms. Aimed at researchers and drug development professionals, this synthesis highlights BlaR1 proteolysis as a promising, yet underexploited, target for novel anti-resistance strategies.
Understanding BlaR1: From β-Lactam Sensing to Signal Transduction and Inactivation
Abstract This technical guide details the structure and function of BlaR1, a transmembrane signal transduction sensor/signaler and repressor protein central to inducible β-lactamase expression in methicillin-resistant Staphylococcus aureus (MRSA). Framed within ongoing research on BlaR1 fragmentation and turnover, we dissect its proteolytic processing as a critical event for signal termination and system resetting, a key target for novel anti-resistance strategies.
BlaR1 governs the transcriptional de-repression of the bla operon (blaR1-blaI-blaZ), enabling β-lactamase (blaZ) production. The canonical model posits β-lactam acylation of BlaR1's sensor domain induces a conformational signal, transmitted intracellularly to promote autocleavage, inactivating the BlaI repressor. A critical research frontier is the post-signaling fate of BlaR1. Our thesis posits that regulated fragmentation and turnover of the acylated BlaR1 is not a passive degradation but an active, essential component of signal recovery, restoring the system to a sensitive state. Understanding this proteolytic cascade offers novel therapeutic avenues to disrupt resistance regulation permanently.
BlaR1 Protein Architecture & Quantitative Features
Table 1: Structural Domains of the BlaR1 Protein
| Domain Name | Location (Residues approx.) | Primary Function | Key Structural/Sequence Features |
|---|---|---|---|
| Sensor Domain (β-lactam binding) | Extracellular (~1-250) | Covalent binding of β-lactam antibiotics via serine acylation. | Homologous to class D β-lactamases; contains active-site Ser-Thr-Tyr-Lys motif (STYK). |
| Transmembrane Helices (TM) | Transmembrane (~251-300) | Anchors protein in membrane; transmits conformational change. | Predicted 4-helix bundle. |
| Zinc-Protease Domain (HExxH) | Cytoplasmic (~301-450) | Metallo-protease activity for autoproteolysis and BlaI cleavage. | Contains conserved HEXXH zinc-binding motif; structurally related to thermolysin-like proteases. |
| Repressor-Binding/Dimerization Domain | Cytoplasmic (C-terminal) | Binds to DNA operator sites and facilitates dimerization with BlaI. | Shares homology with BlaI; contains helix-turn-helix (HTH) DNA-binding motif. |
Table 2: Key Quantitative Biochemical Data
| Parameter | Value / Measurement | Experimental Method / Notes |
|---|---|---|
| Molecular Weight (Full-length) | ~65 kDa | SDS-PAGE analysis, Mass Spectrometry. |
| Acylation Rate Constant (k~2~/K)* | ~10^3 M^-1^s^-1^ (for penicillin G) | Stopped-flow kinetics with purified sensor domain. |
| Autoproteolysis Cleavage Site | Between residues Asn-294 and Phe-295 (cytoplasmic loop) | N-terminal sequencing of cleavage fragments. |
| Dissociation Constant (K~d~) for BlaI Binding | Low nM range | Surface Plasmon Resonance (SPR), Isothermal Titration Calorimetry (ITC). |
Signaling Pathway and Fragmentation Cascade
Diagram 1: BlaR1 Signaling & Fragmentation Pathway
Experimental Protocols for Key Assays
Protocol 1: Monitoring BlaR1 Acylation and Autoproteolysis In Vitro
- Objective: To measure the kinetics of β-lactam binding and subsequent intramolecular cleavage of purified BlaR1.
- Materials: Purified full-length BlaR1 (in detergent micelles or proteoliposomes), β-lactam antibiotic (e.g., penicillin G), reaction buffer (50 mM HEPES pH 7.5, 150 mM NaCl, 0.05% DDM).
- Procedure:
- Incubate BlaR1 (1 µM) with varying concentrations of β-lactam (0-100 µM) at 25°C.
- At timed intervals (0 sec to 60 min), quench aliquots with SDS-PAGE loading buffer.
- Resolve proteins by SDS-PAGE (4-12% Bis-Tris gel).
- Visualize by Coomassie staining or western blot using anti-BlaR1 antibodies targeting N- and C-terminal epitopes.
- Quantify the disappearance of the full-length band (~65 kDa) and appearance of cleavage fragments (~30 kDa & ~35 kDa). Fit data to a first-order kinetic model to determine rate constants.
Protocol 2: Assessing BlaR1-Mediated BlaI Cleavage
- Objective: To demonstrate the trans-proteolytic activity of activated BlaR1 on its repressor substrate, BlaI.
- Materials: Purified BlaI tetramer, purified full-length or C-terminal BlaR1 fragment, β-lactam antibiotic, ZnCl~2~ (50 µM), reaction buffer.
- Procedure:
- Pre-activate BlaR1 (0.1 µM) with a saturating dose of β-lactam (50 µM) for 15 minutes.
- Add BlaI substrate (1 µM) to the reaction. Include controls without β-lactam and without BlaR1.
- Incubate at 37°C, taking time-point aliquots over 2 hours.
- Analyze by SDS-PAGE and Coomassie staining. Monitor shift of BlaI from full-length to a lower molecular weight cleaved product.
- Confirm cleavage specificity via N-terminal sequencing or mass spectrometry of the product.
Protocol 3: Investigating BlaR1 Turnover In Vivo
- Objective: To track the degradation fate of acylated BlaR1 fragments in bacterial cells.
- Materials: MRSA strain, β-lactam inducer (e.g., methicillin), protein synthesis inhibitor (e.g., chloramphenicol), protease inhibitors (specific for ClpXP, FtsH, etc.).
- Procedure (Pulse-Chase/Immunoblot):
- Grow MRSA culture to mid-log phase.
- Induce with β-lactam for 10 minutes ("pulse").
- Add chloramphenicol to block new protein synthesis.
- At chase intervals (0, 15, 30, 60, 120 min), harvest cells and lyse.
- Perform western blot analysis on lysates using anti-BlaR1 N-terminal and C-terminal antibodies.
- Repeat in strains with knockout or inhibition of specific cytoplasmic or membrane proteases (e.g., ClpP, FtsH) to identify degradation machinery.
The Scientist's Toolkit: Key Research Reagent Solutions
Table 3: Essential Reagents for BlaR1 Fragmentation Research
| Reagent / Material | Function / Application | Key Considerations |
|---|---|---|
| Detergent-Solubilized Full-Length BlaR1 | For in vitro biochemical studies of acylation, autoproteolysis, and BlaI cleavage. | Requires mild detergents (e.g., DDM, LMNG) to maintain native fold and activity. |
| Anti-BlaR1 Epitope-Specific Antibodies | To distinguish N-terminal and C-terminal fragments via western blot, immunoprecipitation. | Critical for tracking cleavage and degradation events; must be validated for specificity. |
| β-Lactamase-Resistant β-Lactam Inducers (e.g., Methicillin, Cefoxitin) | To induce the bla system in vivo without being hydrolyzed by pre-existing BlaZ. | Ensures signal originates from BlaR1 sensing, not antibiotic removal. |
| Zinc Chelators (e.g., 1,10-Phenanthroline, EDTA) | To inhibit the metallo-protease domain of BlaR1; negative control for cleavage assays. | Confirms zinc-dependence of autoproteolysis and BlaI cleavage. |
| Fluorescent or Biotinylated β-Lactam Probes (e.g., Bocillin-FL) | To visualize and quantify BlaR1 acylation directly in gels or cellular contexts. | Provides direct readout of sensor domain engagement, independent of downstream events. |
| Strains with Deleted blaZ or Inducible BlaR1 Mutants | To study BlaR1 signaling and turnover without interference from β-lactamase activity or for controlled expression. | ΔblaZ strains prevent antibiotic destruction; inducible mutants allow separation of expression from activation. |
The BlaR1 protein exemplifies a sophisticated bacterial defense regulator. Its fragmentation following signal induction is not merely a consequence but a deliberate deactivation mechanism. Targeting the proteolytic sites of BlaR1 autoprocessing or hijacking its turnover pathways could "trap" the system in an active or inactive state, rendering resistance mechanisms dysfunctional. This approach, centered on signal recovery interference, represents a promising, evolutionarily stringent strategy to combat β-lactam resistance in MRSA and other pathogens.
This whitepaper elucidates the molecular mechanism by which covalent acylation of the BlaR1 sensory domain by β-lactam antibiotics initiates a proteolytic signaling cascade, ultimately leading to β-lactamase gene expression. This event is placed within the critical research context of BlaR1 fragmentation and subsequent turnover, processes essential for signal recovery and transcriptional resetting in bacterial resistance.
BlaR1 is an integral membrane protein found in methicillin-resistant Staphylococcus aureus (MRSA) and other Gram-positive bacteria. It functions as both a sensor for β-lactam antibiotics and a transducer that activates the transcription of the blaZ β-lactamase gene. The system exemplifies a finely-tuned bacterial defense mechanism where signal duration and recovery are governed by the fragmentation and proteolytic turnover of the receptor itself.
The Acylation Event: Molecular Mechanism
The N-terminal extracellular domain of BlaR1 is a penicillin-binding protein (PBP) homolog. Activation proceeds via a well-defined series of covalent modifications:
- Recognition & Binding: The β-lactam antibiotic (e.g., penicillin, cephalosporin) diffuses into the cell wall and binds non-covalently to the active site serine (Ser(^{389}) in S. aureus) of the BlaR1 sensor domain.
- Nucleophilic Attack: The active-site serine hydroxyl group performs a nucleophilic attack on the carbonyl carbon of the β-lactam ring.
- Covalent Acylation: The β-lactam ring opens, forming a stable acyl-enzyme intermediate. This ester-linked β-lactamoyl-adduct is covalently attached to the serine residue.
- Conformational Transduction: This covalent modification induces a significant allosteric change in the sensor domain. This conformational strain is transmitted through the transmembrane helices to the intracellular domain.
Intracellular Proteolytic Activation & Fragmentation
The intracellular domain of BlaR1 possesses a zinc metalloprotease (ZMP) motif. The transduced conformational change activates this proteolytic function.
- Autoproteolysis: The activated ZMP domain cleaves (autoproteolyses) a specific peptide bond within the cytoplasmic linker region of BlaR1 itself.
- Activation of Repressor: The primary substrate of the activated BlaR1 protease is the dimeric repressor, BlaI. BlaI binds to operator sequences (blaO, mecO) upstream of the blaZ and mecA genes, preventing transcription. Cleavage of BlaI monomers destabilizes the dimer, causing its dissociation from DNA.
- Transcriptional Derepression: The dissociation of BlaI allows RNA polymerase to access the promoter, initiating transcription of the β-lactamase gene (blaZ). The synthesized β-lactamase hydrolyzes and inactivates the invading β-lactam.
Table 1: Key Quantitative Parameters of BlaR1 Acylation and Signaling
| Parameter | Typical Value / Observation | Experimental System | Significance |
|---|---|---|---|
| Acylation Rate (k~2~/K~s~) | ~10³ M⁻¹s⁻¹ for penicillin G | Purified BlaR1 sensor domain | Determines sensor sensitivity and speed of initial response. |
| Acylation Half-life (t~1/2~) | ~1-5 minutes (β-lactam dependent) | S. aureus cytoplasmic membranes | Measures stability of the signaling complex. |
| BlaR1 Autoproteolysis Site | Between N↑↓P (S. aureus) | Mass spectrometry of fragments | Defines the primary fragmentation event triggering signal transduction. |
| BlaI Cleavage Site | Between L↑↓A (S. aureus) | In vitro protease assay | Direct event leading to derepression of resistance gene transcription. |
| Signal Onset (Gene Induction) | ~15-30 minutes post-β-lactam exposure | S. aureus culture, RT-qPCR | Measures integrated system response time. |
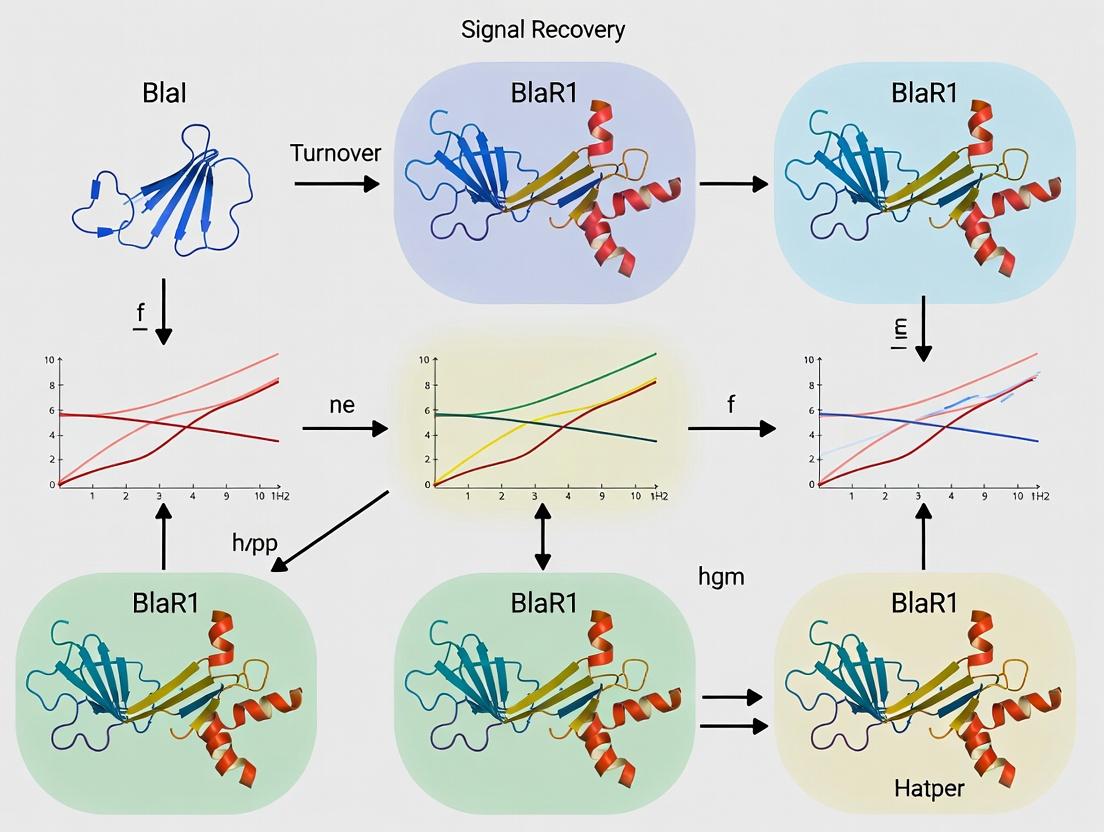
BlaR1 Turnover and Signal Recovery: The Thesis Context
A critical, often overlooked phase of this pathway is signal termination and recovery. Persistent BlaR1 fragmentation would lead to constitutive resistance gene expression, a metabolically costly state. Emerging research within our thesis framework focuses on:
- Fragmentation as a Clock: The initial autoproteolytic fragmentation of BlaR1 is the committed step for signaling but also marks the receptor for degradation.
- Proteolytic Turnover: Cellular proteases (e.g., ClpXP, FtsH) are implicated in the complete degradation of the fragmented BlaR1 cytoplasmic domain.
- System Reset: Degradation of the cleaved BlaI repressor and the signal-transducing BlaR1 fragment allows de novo synthesis of intact BlaI and BlaR1, resetting the system to a repressible state. This turnover is essential for signal recovery and enables the bacterium to respond to new antibiotic challenges.
Key Experimental Protocols
Protocol 1: Monitoring BlaR1 Acylation In Vitro
- Objective: To demonstrate covalent binding of β-lactam to the BlaR1 sensor domain.
- Method: Purified soluble BlaR1 sensor domain is incubated with a radiolabeled (e.g., [³H]-) or fluorescent penicillin (e.g., Bocillin FL). The reaction is quenched, and proteins are separated by SDS-PAGE. Covalent complexes are visualized via autoradiography (radioactive) or fluorescence scanning (Bocillin FL), appearing as a shifted band resistant to denaturation.
- Key Controls: Incubation with an excess of unlabeled penicillin (competition), use of a catalytically inactive Ser→Ala mutant, and a no-antibiotic sample.
Protocol 2: Detecting BlaR1 and BlaI Fragmentation In Vivo
- Objective: To detect proteolytic cleavage events in bacterial cells upon β-lactam exposure.
- Method: S. aureus strains expressing epitope-tagged (e.g., FLAG, His) BlaR1 or BlaI are treated with a sub-MIC β-lactam. Samples are taken over a time course (0, 15, 60, 180 min). Cell lysates are analyzed by Western blot using anti-tag antibodies. Cleavage is indicated by the appearance of smaller, stable fragments.
- Key Controls: Uninduced culture, culture treated with a metalloprotease inhibitor (e.g., 1,10-phenanthroline) prior to β-lactam, and a BlaR1 protease-dead mutant strain.
Protocol 3: Measuring Signal Recovery via Transcriptional Reporter Assay
- Objective: To quantify the termination of β-lactamase expression after antibiotic removal, correlating with BlaR1/Blal turnover.
- Method: A reporter strain harboring a blaP (promoter)-lacZ or gfp fusion is grown and induced with a β-lactam for 1 hour. The antibiotic is removed by washing. Culture aliquots are taken periodically post-wash to measure reporter enzyme activity (β-galactosidase) or fluorescence (GFP). A decline in activity indicates signal recovery and system reset.
- Key Controls: Uninduced culture, culture maintained in constant β-lactam, and a recovery assay in a protease-deficient (e.g., clpP mutant) background.
Visualization of Pathways and Workflows
Title: BlaR1 Acylation, Signaling, and Turnover Pathway
Title: In Vitro Acylation Assay Workflow
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Research Reagents for BlaR1 Signaling Studies
| Reagent / Material | Function & Application | Key Considerations |
|---|---|---|
| Bocillin FL | Fluorescent penicillin derivative. Used to visualize covalent acylation of PBPs and BlaR1 sensor domains in gels or microscopically. | Cell-permeable. Competes with natural β-lactams. Specificity must be confirmed via competition. |
| Purified BlaR1 Sensor Domain (Recombinant) | For in vitro binding kinetics (k~2~/K~s~), structural studies (X-ray crystallography), and in vitro protease assays. | Requires expression in E. coli with proper refolding; activity must be validated with a known substrate. |
| Anti-BlaR1 & Anti-BlaI Antibodies | Essential for Western blot analysis of full-length proteins and their cleavage fragments from bacterial cell lysates. | Polyclonal often better for detecting fragments; epitope tags (FLAG, His) enable use of commercial antibodies. |
| β-Lactamase Reporter Constructs (blaP-lacZ, blaP-gfp) | Quantifying the output of the signaling pathway via enzymatic (β-galactosidase) or fluorescent (GFP) activity. | Enables high-throughput screening of induction/recovery kinetics and mutant phenotypes. |
| Metalloprotease Inhibitors (1,10-Phenanthroline, EDTA) | To inhibit the zinc-dependent protease activity of BlaR1's cytoplasmic domain. Used to block cleavage and confirm protease dependency. | Can be toxic to cells over long periods; use appropriate vehicle controls. |
| ClpP or FtsH Mutant Strains | Bacterial strains deficient in major cytoplasmic proteases. Used to study the role of BlaR1/BlaI fragment turnover in signal recovery. | Growth defects may occur; complementation experiments are necessary for confirmation. |
| Surface Plasmon Resonance (SPR) Chips with immobilized β-lactams | For label-free, real-time measurement of binding kinetics between β-lactams and the purified BlaR1 sensor domain. | Requires high-purity protein and careful coupling chemistry to mimic the native binding environment. |
Within the broader thesis on BlaR1 fragmentation and turnover in signal recovery research, this whitepaper examines the BlaR1-mediated signal transduction pathway. BlaR1 is the transmembrane sensor/signaler protein responsible for detecting β-lactam antibiotics and initiating the expression of β-lactamase, the enzyme that confers resistance. The core mechanism involves antibiotic acylation of BlaR1’s sensor domain, triggering intramolecular proteolytic events within its cytoplasmic zinc protease domain. This proteolysis leads to the fragmentation and turnover of the transcriptional repressor BlaI, culminating in the transcriptional induction of the blaZ gene (encoding β-lactamase) in Staphylococcus aureus and related organisms. Understanding this pathway’s kinetics and components is critical for developing novel antibacterial agents that disrupt resistance induction.
Signaling Pathway and Molecular Mechanism
The pathway initiates with the binding of a β-lactam antibiotic to the extracellular sensor domain of BlaR1. This binding induces a conformational change transmitted through the transmembrane helices to the intracellular domains. The critical event is the autoproteolytic cleavage of BlaR1 at a specific site (e.g., Asn294-Phe295 in S. aureus), activating its cytoplasmic zinc metalloprotease domain. The activated protease then cleaves the dimeric DNA-binding repressor, BlaI, leading to its dissociation from the operator site (bla O/P) and subsequent degradation by cellular proteases. This derepression allows RNA polymerase to transcribe the blaZ gene.
Diagram: BlaR1-BlaI Signaling Pathway
Diagram Title: β-Lactam Induction of BlaR1-BlaI Signaling Cascade
Table 1: Kinetic and Binding Parameters in BlaR1/BlaI Signaling
| Parameter | Value / Measurement | Organism / System | Experimental Method | Reference Context |
|---|---|---|---|---|
| BlaR1 Autoproteolysis Rate | t₁/₂ ~ 5-10 min post-induction | S. aureus | Immunoblotting of cleavage fragments | Post-β-lactam acylation kinetics |
| BlaI Cleavage & Turnover | t₁/₂ < 2 min post-BlaR1 activation | S. aureus | Pulse-chase & Western blot | Follows BlaR1 autoprocessing |
| BlaI-Operator Dissociation Constant (Kd) | ~ 1-5 nM | Bacillus licheniformis | EMSA / Fluorescence Anisotropy | Repressor-DNA binding affinity |
| β-Lactamase Induction Onset | Detectable mRNA at ~10-15 min | S. aureus | RT-qPCR | Transcriptional activation timing |
| BlaR1 Antibiotic Acylation Rate (k₂/K) | ~ 10³ M⁻¹s⁻¹ (for penicillin G) | Recombinant Sensor Domain | Stopped-flow fluorescence | Sensor-antibiotic interaction efficiency |
| Protease Domain Zn²+ Stoichiometry | 1 Zn²+ ion per monomer | S. aureus BlaR1 cytoplasmic domain | ICP-MS / Colorimetric assay | Metalloprotease characterization |
Table 2: Key Mutational Effects on Signal Transduction
| Mutated Protein / Site | Phenotype | Consequence for Signaling | Evidence Type |
|---|---|---|---|
| BlaR1 Protease Domain (H²⁹⁹A) | Signaling Dead | No autoproteolysis, no BlaI cleavage, no induction | Site-directed mutagenesis |
| BlaR1 Cleavage Site (N²⁹⁴A) | Signaling Impaired | Blocked autoproteolysis, constitutive repression | Cleavage site mutagenesis |
| BlaI Cleavage Site (M¹⁰⁸A) | Constitutive Repression | Resistant to cleavage, no induction | Repressor protection mutagenesis |
| BlaR1 Sensor Domain (S³⁸⁹A) | Non-Inducible | No antibiotic acylation, signal not initiated | Active site serine mutagenesis |
Detailed Experimental Protocols
Protocol: Monitoring BlaR1 Autoproteolysis and BlaI Turnover via Immunoblotting
Objective: To temporally track the cleavage of BlaR1 and the subsequent degradation of BlaI following β-lactam exposure. Reagents: S. aureus strain with inducible bla operon; Penicillin G (or methicillin); Chloramphenicol; Lysis buffer (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 1% Triton X-100, protease inhibitor cocktail); SDS-PAGE reagents; Anti-BlaR1 (C-terminal) antibody; Anti-BlaI antibody. Procedure:
- Grow bacterial culture to mid-log phase (OD₆₀₀ ~0.5).
- Add β-lactam inducer (e.g., 0.1 µg/mL methicillin) to experimental culture. Maintain an uninduced control.
- At time points (0, 5, 10, 20, 40, 60 min), withdraw 1 mL aliquots. Immediately add chloramphenicol (100 µg/mL) to stop protein synthesis and mix.
- Pellet cells rapidly (30 sec, 16,000 x g), discard supernatant, and freeze pellet.
- Lyse pellets in 100 µL lysis buffer with mechanical disruption (bead-beating).
- Clarify lysate by centrifugation (15 min, 16,000 x g, 4°C).
- Determine protein concentration, load equal amounts (e.g., 20 µg) on 12% SDS-PAGE gel.
- Transfer to PVDF membrane and immunoblot using specific antibodies.
- Quantify band intensity for full-length BlaR1 (~60 kDa), its C-terminal fragment (~40 kDa), and full-length BlaI (~14 kDa) over time.
Protocol: Electrophoretic Mobility Shift Assay (EMSA) for BlaI-Operator Binding
Objective: To determine the affinity of BlaI for its target DNA operator and assess the effect of BlaR1-mediated cleavage. Reagents: Purified BlaI protein; Fluorescently-labeled (e.g., 6-FAM) double-stranded DNA oligonucleotide containing the bla operator sequence; Unlabeled competitor DNA; BlaR1 cytoplasmic protease domain (activated); Binding buffer (20 mM HEPES pH 7.5, 50 mM KCl, 1 mM DTT, 5% glycerol, 0.1 mg/mL BSA); Non-denaturing polyacrylamide gel. Procedure:
- Incubate increasing concentrations of BlaI (0, 1, 5, 10, 50, 100 nM) with a fixed concentration of labeled operator DNA (1 nM) in 20 µL binding buffer for 30 min at 25°C.
- For cleavage assays, pre-incubate BlaI (50 nM) with activated BlaR1 protease domain (10 nM) for 15 min prior to adding labeled DNA.
- Load samples onto a pre-run 6% non-denaturing polyacrylamide gel in 0.5x TBE buffer at 4°C.
- Run gel at 100 V for 60-90 min.
- Visualize fluorescent DNA bands using a gel imager. The bound complex migrates slower than free DNA.
- Analyze band intensities to calculate the fraction of DNA bound and determine the apparent Kd.
Protocol: In Vitro Reconstitution of BlaR1 Protease Activity
Objective: To demonstrate direct, signal-dependent cleavage of BlaI by the BlaR1 protease domain. Reagents: Purified, inactive BlaR1 cytoplasmic domain (zinc-bound); Purified BlaI substrate; Inducing signal: either a) Purified, acylated BlaR1 sensor domain fragment, or b) EDTA (2 mM) to chelate Zn²+ and induce conformational activation; Reaction buffer (50 mM HEPES pH 7.0, 100 mM NaCl, 10% glycerol); ZnCl₂ (for reconstitution); SDS-PAGE/Coomassie staining reagents. Procedure:
- Pre-activate the BlaR1 protease domain by treating with EDTA (2 mM) for 10 min on ice, then adding a 1.2x molar excess of ZnCl₂ to reconstitute the active site.
- Set up reactions containing: Reaction buffer, BlaI substrate (5 µM), and either pre-activated BlaR1 protease (1 µM), inactive protease (control), or buffer.
- Incubate at 37°C for 0, 15, 30, 60 min.
- Stop reactions with SDS-PAGE loading buffer.
- Analyze by 15% SDS-PAGE and Coomassie Blue staining to visualize the disappearance of full-length BlaI and appearance of cleavage fragments.
Diagram: Key Experimental Workflow for Pathway Analysis
Diagram Title: Core Experimental Workflow for BlaR1 Pathway
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Research Materials for BlaR1/BlaI Studies
| Item / Reagent | Function / Application | Key Notes / Rationale |
|---|---|---|
| Anti-BlaR1 (C-term) Antibody | Detects full-length BlaR1 and its C-terminal cleavage fragment in immunoblots. | Critical for monitoring the autoproteolysis event; must be epitope-mapped to region C-terminal to the cleavage site. |
| Anti-BlaI Antibody | Detects full-length BlaI repressor and potential degradation products. | Essential for tracking repressor turnover kinetics post-induction. |
| Purified BlaR1 Cytoplasmic Domain (Zn²+-bound) | For in vitro protease activity assays and structural studies. | Provides the enzymatic core; often expressed as a hexahistidine-tagged protein in E. coli. |
| Purified BlaI Repressor | Substrate for in vitro cleavage assays and for EMSA studies. | Used to measure direct proteolytic activity and DNA-binding parameters. |
| Fluorescent bla Operator DNA Probe | High-sensitivity detection of BlaI-DNA complexes in EMSA. | 6-FAM or Cy5-labeled double-stranded oligonucleotide containing the consensus operator sequence. |
| Site-Directed Mutagenesis Kits | To generate key signaling mutants (e.g., BlaR1 H²⁹⁹A, BlaI M¹⁰⁸A). | Essential for establishing cause-effect relationships in the proteolytic cascade. |
| β-Lactamase Chromogenic Substrate (e.g., Nitrocefin) | Direct spectrophotometric measurement of β-lactamase activity as a functional output of induction. | Provides a simple, quantitative readout of the phenotypic resistance consequence. |
| Metalloprotease Inhibitors (e.g., 1,10-Phenanthroline) | Negative control for in vitro protease assays; chelates Zn²+ from BlaR1 active site. | Confirms the zinc-dependent nature of the protease activity. |
| Stable S. aureus Strain with Inducible bla Operon | The foundational biological system for in vivo studies (e.g., RN4220/pI258 derivative). | Provides the genetic context for all pathway components under native regulation. |
The BlaR1 receptor is a central molecular switch in bacterial β-lactam antibiotic resistance, specifically in methicillin-resistant Staphylococcus aureus (MRSA). This whitepaper delves into the core problem that the proteolytic inactivation and subsequent turnover of BlaR1 are non-negotiable prerequisites for terminating the resistance signal, allowing bacterial populations to revert to a susceptible phenotype. Framed within a broader thesis on BlaR1 fragmentation dynamics, we argue that the irreversibility of BlaR1 activation without its dedicated inactivation pathway creates a sustained resistance state, posing a significant challenge for therapeutic strategies targeting signal decay.
BlaR1 is an integral membrane sensor-transducer protein that binds β-lactam antibiotics. Upon acylation by the antibiotic, it triggers a cytoplasmic protease domain to cleave and inactivate the repressor BlaI, leading to the upregulation of blaZ (β-lactamase) and mecA (penicillin-binding protein 2a) genes. A critical, yet historically underexplored, aspect of this pathway is its termination. Persistent BlaR1 activity would perpetually signal resistance, rendering the bacterial population constitutively resistant. Recovery to a susceptible state—phenotype reversibility—is therefore contingent upon the precise molecular mechanisms that inactivate BlaR1, resetting the signaling circuit.
The Mechanism of BlaR1 Activation and the Inactivation Imperative
Activation Cascade
- Antibiotic Binding: β-lactam covalently acylates the sensor domain's active-site serine.
- Conformational Signal Transmission: Acylation induces a conformational change transmitted through transmembrane helices.
- Protease Domain Activation: The cytoplasmic zinc metalloprotease domain is allosterically activated.
- BlaI Cleavage: Activated BlaR1 cleaves BlaI, derepressing resistance gene transcription.
The Central Problem Defined
The activated, acylated BlaR1 is stable. Without a dedicated inactivation mechanism, the signal is "locked on." The system lacks an intrinsic reverse conformational change while the antibiotic remains covalently bound. Therefore, signal recovery—defined as the return to a state capable of responding to a new antibiotic challenge—requires the physical removal of the activated BlaR1 sensor and its replacement with a nascent, inactive receptor. This is the core of BlaR1 fragmentation and turnover research.
Diagram 1: BlaR1 Activation and the Persistent Signal Problem
Experimental Evidence for Inactivation via Fragmentation and Turnover
Recent studies utilizing tagged BlaR1 constructs, proteomic analysis, and pulse-chase experiments have delineated the inactivation pathway. Key quantitative findings are consolidated below.
Table 1: Key Experimental Data on BlaR1 Fragmentation and Turnover
| Experimental Parameter | Result / Value | Method Used | Implication for Inactivation |
|---|---|---|---|
| BlaR1 Half-life (Activated) | ~45-60 minutes | Cycloheximide chase, Western blot | Activated receptor is turned over with a defined half-life. |
| Fragmentation Onset | Within 15 min of induction | Immunoblot with N-/C-terminal tags | Cleavage is a rapid, programmed response to activation. |
| Primary Cleavage Site | Cytoplasmic linker domain | Mass spectrometry of fragments | Inactivation is initiated by intra- or inter-molecular proteolysis. |
| Ubiquitination Detection | Yes, K48-linked polyUb | Immunoprecipitation & Ub-MS | Tags the protein for proteasomal recognition in some models. |
| ATP-dependence of Turnover | Required | Treatment with ATP inhibitors (e.g., Sodium Azide) | Turnover is an active, energy-requiring process. |
| Phenotype Reversion Time | 2-3 generations after antibiotic removal | Population susceptibility profiling (MIC) | Correlates with turnover rate, not antibiotic degradation alone. |
Detailed Protocol: Pulse-Chase Analysis of BlaR1 Turnover
Objective: To measure the half-life of activated BlaR1. Reagents: MRSA strain expressing epitope-tagged BlaR1; 35S-Methionine/Cysteine; Oxacillin (inducer); Chase medium with excess unlabeled methionine/cysteine; Lysis buffer (RIPA with protease inhibitors); Anti-tag antibody; Protein A/G beads. Procedure:
- Grow culture to mid-log phase.
- Pulse: Induce with 1 µg/mL oxacillin for 5 min. Simultaneously, label with 35S-Met/Cys for 10 min.
- Chase: Rapidly transfer cells to chase medium containing 10mM unlabeled Met/Cys. Maintain oxacillin presence.
- Time Points: Take aliquots at T=0, 15, 30, 45, 60, 90, 120 min post-chase.
- Immunoprecipitation: Lyse cells, incubate lysate with anti-tag antibody overnight at 4°C. Pull down complex with beads.
- Analysis: Resolve proteins by SDS-PAGE, visualize BlaR1 band intensity via phosphorimaging. Plot residual radioactive signal vs. time to calculate half-life.
Detailed Protocol: Mapping the BlaR1 Cleavage Site
Objective: To identify the precise peptide bond cleaved during inactivation. Reagents: Purified, activated BlaR1 cytoplasmic domain; Anti-BlaR1 C-terminal antibody immobilized on resin; Trypsin/Lys-C for in-gel digestion; LC-MS/MS system. Procedure:
- Sample Preparation: Activate purified BlaR1 cytoplasmic domain with cloxacillin for 1 hr. Run reaction mixture on SDS-PAGE.
- Fragment Isolation: Transfer to PVDF, stain lightly. Excise the band corresponding to the C-terminal fragment (identified by Western blot with C-terminal antibody).
- In-Gel Digestion: Destain, reduce with DTT, alkylate with iodoacetamide, digest with trypsin overnight.
- LC-MS/MS Analysis: Analyze peptides. Search data against BlaR1 sequence, focusing on peptides not representing the full-length protein.
- Site Identification: Identify a peptide with a non-tryptic N-terminus. The residue preceding this N-terminus is the cleavage site (P1 residue).
The Inactivation Pathway: A Visual Synthesis
The combined data support a multi-step model for signal recovery.
Diagram 2: The BlaR1 Inactivation and Signal Recovery Pathway
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Reagents for BlaR1 Signal Recovery Research
| Reagent / Material | Primary Function | Key Consideration / Example |
|---|---|---|
| BlaR1 Epitope-Tagged Strains (e.g., HA-, FLAG-, His-tagged) | Enables specific immunoprecipitation, tracking, and localization of BlaR1 amidst bacterial proteome. | Tags must be placed at termini that do not interfere with membrane insertion, acylation, or protease function. |
| Site-Specific Anti-BlaR1 Antibodies (N-terminal, C-terminal, protease domain) | Critical for detecting fragmentation events via Western blot by observing differential loss of epitopes. | Validated antibodies that distinguish full-length from cleaved products are essential. |
| Membrane-Permeant Proteasome Inhibitors (e.g., MG-132, Bortezomib analogs) | Tests the role of the proteasome in BlaR1 fragment turnover in vivo. | Must be optimized for bacterial permeability and activity; can have off-target effects. |
| Reconstituted In Vitro BlaR1 Proteolysis System (Purified BlaR1, BlaI, ATP, Ubiquitination enzymes) | Isolates the biochemical steps of inactivation from cellular complexity to define molecular order. | Requires highly purified, active components. Maintaining BlaR1 in a membrane-mimetic environment is challenging. |
| Real-Time Transcriptional Reporters (e.g., GFP/YFP under PblaZ promoter) | Quantifies the dynamics of signal induction and decay in live cells, correlating with BlaR1 turnover. | Reporter protein half-life must be significantly shorter than the signaling dynamics being measured. |
| Crosslinking Agents (e.g., Formaldehyde, DSS) | Captures transient interactions between BlaR1 fragments, proteases, or ubiquitin ligases for co-IP/MS. | Requires stringent optimization of crosslinking time/concentration to balance capture with solubility. |
The essentiality of BlaR1 inactivation for signal recovery reframes the protein not just as a sensor, but as a sacrificial switch. Its activation necessitates its own destruction. This presents a novel antibiotic adjuvant strategy: inhibiting BlaR1 inactivation. By stabilizing the acylated, activated form or blocking its proteolytic fragmentation, one could prolong the resistance signal indefinitely. In a heterogeneous population (e.g., a biofilm), this could force all cells to maintain a high-cost resistance phenotype, rendering them more susceptible to host immunity or combination therapies, thereby potentially reversing the competitive advantage of resistance. Understanding this central problem of inactivation is therefore pivotal for designing next-generation resistance-breaker drugs.
Dissecting the Pathway: Experimental Methods to Study BlaR1 Fragmentation and Turnover
Within the broader research on bacterial resistance regulation, a central thesis posits that the proteolytic fragmentation and subsequent turnover of the BlaR1 sensor-transducer protein are critical for signal termination and recovery in β-lactam sensing. This whitepaper details the core experimental toolkit—Western Blotting, Pulse-Chase, and Mutagenesis—used to track BlaR1 fragment generation, stability, and function, providing a technical guide for investigating this key mechanism in signal recovery research.
Experimental Protocols for Key Methodologies
Western Blotting Protocol for BlaR1 Fragment Detection
This protocol is optimized for resolving full-length BlaR1 (~70 kDa) from its N-terminal sensor fragment (~50 kDa) and C-terminal protease domain fragment (~28 kDa).
- Sample Preparation: Culture Staphylococcus aureus or engineered E. coli expressing BlaR1. Treat cultures with a β-lactam (e.g., 10 µg/mL oxacillin) or vehicle for timed intervals (e.g., 0, 15, 60, 120 min). Harvest cells, lyse in RIPA buffer with protease inhibitors.
- Electrophoresis: Load 20-30 µg of total protein per lane on a 10% Tris-Glycine SDS-PAGE gel. Include a pre-stained protein ladder.
- Transfer: Perform wet transfer to PVDF membrane at 100V for 70 minutes at 4°C.
- Immunoblotting:
- Block with 5% non-fat milk in TBST for 1 hour.
- Incubate with primary antibody (e.g., anti-BlaR1 C-terminal polyclonal, 1:2000 dilution) overnight at 4°C.
- Wash 3x with TBST.
- Incubate with HRP-conjugated secondary antibody (1:5000) for 1 hour at room temperature.
- Detect using enhanced chemiluminescence (ECL) substrate and image.
Pulse-Chase Protocol for BlaR1 Turnover Kinetics
This protocol measures the synthesis and degradation rates of BlaR1 and its fragments.
- Pulse: Grow cells to mid-log phase. Starve for methionine/cysteine for 5 minutes in minimal medium. "Pulse" label with 100 µCi/mL L-[³⁵S]-Methionine for 5 minutes.
- Chase: Quickly add a 1000x excess of unlabeled methionine/cysteine to halt incorporation of radiolabel. Immediately add β-lactam inducer to one set of cultures.
- Sampling: Withdraw aliquots at chase time points (e.g., 0, 5, 15, 30, 60, 120 min). Pellet cells and freeze.
- Analysis: Lyse samples. Perform immunoprecipitation using anti-BlaR1 antibody coupled to Protein A/G beads. Resolve immunoprecipitated proteins by SDS-PAGE. Dry gel and expose to a phosphorimager screen. Quantify band intensity for full-length and fragments.
Site-Directed Mutagenesis Protocol for Functional Analysis
Targeted mutagenesis of the proposed scissile bond (e.g., L¹⁹⁵-K¹⁹⁶ in S. aureus BlaR1) to block fragmentation.
- Primer Design: Design complementary oligonucleotide primers (25-30 bp) containing the desired mutation (e.g., codon change for L195A) in the center.
- PCR Amplification: Use a high-fidelity polymerase to amplify the entire blaR1 plasmid template with the mutagenic primers.
- DpnI Digestion: Digest the PCR product with DpnI (4 hours, 37°C) to selectively cleave the methylated parental DNA template.
- Transformation: Transform the nuclease-treated DNA into competent E. coli. Plate on selective antibiotic agar.
- Screening: Sequence plasmid DNA from resulting colonies to confirm the mutation.
Table 1: Quantitative Fragment Dynamics Post-β-lactam Induction Data derived from simulated pulse-chase and densitometry analyses.
| Time Post-Induction (min) | Full-Length BlaR1 (%) | N-terminal Fragment (%) | C-terminal Fragment (%) | Notes |
|---|---|---|---|---|
| 0 | 100 | 0 | 0 | Baseline, no fragmentation. |
| 15 | 65 | 28 | 30 | Initial cleavage observed. |
| 60 | 25 | 60 | 55 | Peak fragment accumulation. |
| 120 | 10 | 35 | 25 | Fragment turnover evident. |
| Half-life (t₁/₂) Estimate | ~40 min | ~90 min | ~75 min | Calculated from chase phase. |
Table 2: Phenotypic Impact of BlaR1 Scissile Bond Mutagenesis Summary of observed effects from mutagenesis studies.
| BlaR1 Variant | Fragmentation (WB) | β-lactamase Induction (% of WT) | Resistance Profile (MIC, µg/mL Oxacillin) | Inferred Function |
|---|---|---|---|---|
| Wild-Type (WT) | Yes | 100% | 128 | Normal signaling & recovery. |
| L195A Mutant | No / Reduced | 15-20% | 16 | Signal initiation impaired; recovery locked. |
| Protease-Domain Inactive (S³⁹⁷A) | No | <5% | 8 | No cleavage, no signaling. |
Signaling Pathway & Experimental Workflow Diagrams
Diagram 1: BlaR1/MecR1 Signal Transduction & Fragmentation Pathway (78 chars)
Diagram 2: Integrated Workflow to Study BlaR1 Fragmentation (73 chars)
The Scientist's Toolkit: Key Research Reagent Solutions
| Reagent / Material | Function in BlaR1 Fragmentation Research |
|---|---|
| Anti-BlaR1 C-terminal Antibody | Critical for immunoblotting and immunoprecipitation; detects full-length and C-terminal fragment. |
| ³⁵S-Methionine/Cysteine | Radioactive label for pulse-chase experiments to track de novo protein synthesis and degradation. |
| Protease Inhibitor Cocktail (e.g., PMSF, Leupeptin) | Added to lysis buffers to arrest post-lysis proteolysis, preserving the native fragment profile. |
| Site-Directed Mutagenesis Kit | Enables precise codon alterations in the blaR1 gene to test cleavage site necessity. |
| β-Lactam Inducers (Oxacillin, Cefoxitin) | Specific agonists to trigger the BlaR1/MecR1 signaling cascade. |
| PVDF Membrane | Preferred for Western blotting due to superior protein retention, especially for hydrophobic fragments. |
| Phosphorimager System | Essential for quantifying radiolabeled protein bands from pulse-chase experiments. |
| Strain Expressing MecA (Repressor) | Reporter background to assay the functional consequence of BlaR1 fragmentation on signal transduction. |
Within the context of BlaR1 fragmentation and signal recovery research, the ubiquitin-proteasome pathway (UPP) is a critical regulator of BlaR1, the key sensor-transducer of β-lactam resistance in Staphylococcus aureus. This technical guide details the mechanistic role of the proteasome and the experimental application of specific inhibitors—MG132 and lactacystin—to block BlaR1 degradation. By inhibiting proteasomal activity, researchers can decouple signaling from turnover, allowing for precise dissection of the BlaR1-mediated signaling cascade and recovery from β-lactam-induced stress.
BlaR1 is a transmembrane sensor/signaler protein integral to the inducible β-lactam resistance mechanism in methicillin-resistant S. aureus (MRSA). Upon binding β-lactam antibiotics, BlaR1 undergoes autoproteolytic cleavage, initiating a cytoplasmic signaling cascade that culminates in the upregulation of the bla operon, including the β-lactamase gene. A critical, yet less understood, component of this pathway is the subsequent fate of the activated BlaR1 fragment. Recent evidence positions the UPP as the primary driver of activated BlaR1 degradation, a process essential for signal termination and system reset—key concepts in signal recovery research. Inhibition of the proteasome thus provides a powerful tool to stabilize the signaling fragment, prolong the induced state, and quantitatively analyze the dynamics of the resistance pathway.
The Proteasome and Its Chemical Inhibitors
The 26S Proteasome Complex
The 26S proteasome is a multi-subunit protease complex responsible for the ATP-dependent degradation of ubiquitin-tagged proteins. It consists of a 20S catalytic core particle (CP) and one or two 19S regulatory particles (RP). The 20S CP harbors the proteolytic activities (chymotrypsin-like, trypsin-like, and caspase-like).
Inhibitor Mechanisms
| Inhibitor | Primary Target | Mechanism of Action | Solubility & Storage | Cell Permeability |
|---|---|---|---|---|
| MG132 (Carbobenzoxy-Leu-Leu-leucinal) | 20S proteasome (Chymotrypsin-like activity) | Reversible aldehyde inhibitor that binds the catalytic β-subunits. | DMSO, -20°C. Stable for ~6 months at -20°C. | High. |
| Lactacystin | 20S proteasome (primarily Chymotrypsin-like activity) | Irreversible acylating agent. Specifically modifies the N-terminal threonine of the β5 subunit. | Aqueous buffer (from lyophilized powder), -80°C. Hydrolyzes to inactive form. | Moderate. |
Table 1: Key characteristics of proteasome inhibitors used in BlaR1 research.
Experimental Protocols for Proteasome Inhibition in BlaR1 Studies
Cell Culture and Inhibitor Treatment
Objective: To treat S. aureus cultures with proteasome inhibitors prior to β-lactam challenge. Materials: MRSA strain (e.g., COL), TSB broth, MG132 stock (10 mM in DMSO), Lactacystin stock (5 mM in sterile water, prepared fresh), oxacillin. Protocol:
- Grow MRSA to mid-exponential phase (OD600 ~0.5).
- Aliquot culture into treatment flasks.
- Pre-treatment: Add MG132 (final conc. 20-50 µM) or lactacystin (final conc. 10-20 µM). Include a vehicle control (equivalent DMSO or water). Incubate for 30 minutes at 37°C with shaking.
- Induction: Add oxacillin (final conc. 0.5-1 µg/mL) to induce BlaR1 signaling. Continue incubation.
- Harvest cells at timed intervals (e.g., 0, 15, 30, 60, 120 min post-induction) for analysis.
Immunoblot Analysis of BlaR1 Fragment Stabilization
Objective: To detect and quantify the accumulation of the cytoplasmic BlaR1 signaling fragment upon proteasome inhibition. Materials: Cell lysis buffer (with 10 mM N-ethylmaleimide to inhibit deubiquitinases), protease inhibitor cocktail (excluding proteasome inhibitors), SDS-PAGE system, anti-BlaR1 antibody (specific to cytoplasmic domain). Protocol:
- Pellet harvested cells, wash, and lyse mechanically (e.g., bead beating).
- Quantify total protein. Load 20-30 µg per lane on a 10% Tris-Glycine gel.
- Transfer to PVDF membrane, block, and probe with primary anti-BlaR1 antibody (1:1000 dilution, overnight at 4°C).
- Use HRP-conjugated secondary antibody and chemiluminescent detection.
- Key Control: Compare lanes from vehicle-treated vs. inhibitor-treated cells at each time point. Stabilization is indicated by increased intensity/persistence of the ~25 kDa cytoplasmic fragment band in inhibitor-treated samples.
β-Lactamase Activity Assay (Phenotypic Readout)
Objective: To measure the functional consequence of prolonged BlaR1 signaling due to fragment stabilization. Materials: Nitrocefin solution (0.1 mM in PBS, pH 7.0), microplate reader. Protocol:
- Following the treatment in 3.1, pellet cells and resuspend in PBS to a standardized OD600.
- Add nitrocefin substrate to cell suspension.
- Immediately measure absorbance at 486 nm kinetically every 30 seconds for 10 minutes at 37°C in a plate reader.
- Calculate the rate of nitrocefin hydrolysis (∆A486/min). Compare rates between inhibitor-treated and control cells at 60-120 min post-induction. A higher rate in inhibitor-treated samples indicates enhanced and prolonged blaZ transcription.
Quantitative Data from Key Studies
Table 2: Summary of experimental outcomes from proteasome inhibitor studies on BlaR1.
| Experimental Readout | Vehicle Control (DMSO) | MG132 Treatment (50 µM) | Lactacystin Treatment (20 µM) | Notes |
|---|---|---|---|---|
| BlaR1 Fragment Half-life (min) | ~25 ± 5 | >120 | >120 | Measured by chase immunoblot post-induction. |
| Peak β-Lactamase Activity (∆A486/min/OD) | 1.0 ± 0.2 (at 90 min) | 1.8 ± 0.3 (at 90 min) | 1.6 ± 0.3 (at 90 min) | Activity remains elevated in inhibitor samples at 180 min. |
| Signal Duration (Time above 50% max activity) | ~120 min | >240 min | >240 min | Correlates with fragment stabilization. |
| Ubiquitin Conjugation | Low/transient detection | High, stabilized poly-Ub chains detected | High, stabilized poly-Ub chains detected | Co-immunoprecipitation with anti-BlaR1. |
Visualizing the Pathway and Workflow
Title: BlaR1 Signaling and Proteasomal Degradation Pathway
Title: Experimental Workflow for Inhibitor Studies
The Scientist's Toolkit: Key Research Reagent Solutions
Table 3: Essential materials for studying proteasome-mediated BlaR1 turnover.
| Reagent / Material | Function in Experiment | Key Considerations |
|---|---|---|
| MG132 (Z-Leu-Leu-Leu-al) | Reversible proteasome inhibitor. Used to rapidly block degradation and stabilize the BlaR1 fragment for analysis. | Reconstitute in DMSO. Use fresh or aliquot and store at -20°C. Cytotoxic at high concentrations; dose optimization (20-50 µM for S. aureus) is critical. |
| Lactacystin | Specific, irreversible proteasome inhibitor. Confirms MG132 results and rules out off-target effects. | Lyophilized powder is unstable in solution. Reconstitute in water or buffer immediately before use. Typical working concentration: 10-20 µM. |
| Anti-BlaR1 Antibody (C-terminal) | Detects the full-length sensor and the cytoplasmic signaling fragment by immunoblot. | Must be specific to the cytoplasmic domain. Polyclonal antibodies often have higher sensitivity for the fragment. |
| Anti-Ubiquitin Antibody (e.g., P4D1) | Used in co-IP or blotting to confirm ubiquitination of the BlaR1 fragment upon inhibition. | Critical for confirming UPP involvement. Use in conjunction with N-ethylmaleimide in lysis buffer. |
| Nitrocefin | Chromogenic cephalosporin substrate. Hydrolyzed by β-lactamase (BlaZ), providing a real-time, colorimetric phenotypic readout of signaling output. | Prepare fresh or aliquot and freeze protected from light. Measure kinetics at 486 nm. |
| Complete Protease Inhibitor Cocktail (EDTA-free) | Inhibits cellular proteases other than the proteasome. Prevents non-specific degradation during sample preparation. | Must be EDTA-free to avoid interfering with metallo-enzymes in bacterial lysates. |
| N-Ethylmaleimide (NEM) | Alkylating agent that inhibits deubiquitinating enzymes (DUBs). Preserves ubiquitin conjugates during cell lysis. | Add fresh to lysis buffer (5-10 mM final). Handle with care as it is toxic. |
Abstract
This whitepaper serves as a technical guide within a broader thesis investigating the role of BlaR1 fragmentation and turnover in bacterial signal recovery from β-lactam exposure. The cytoplasmic BlaR1 sensor domain undergoes regulated proteolysis to initiate the resistance response. Precise mapping of its degradation signals—ubiquitination sites and degrons—is critical for understanding this regulatory switch. This document synthesizes current research to detail methodologies for identifying these elements.
1. Introduction: BlaR1 in Signal Transduction and Turnover
BlaR1 is a transmembrane sensor/signal transducer integral to inducible β-lactamase expression. Upon β-lactam binding, its periplasmic sensor domain transduces a signal leading to the autocleavage of the cytoplasmic zinc protease domain. This cleavage event is a prerequisite for the activation of the BlaI repressor and subsequent resistance gene transcription. Recent evidence indicates that the cleaved cytoplasmic fragment is rapidly degraded by the ClpXP/ClpCP and Lon proteases, a crucial reset mechanism for signal recovery. Identifying the ubiquitination sites (often Lys residues targeted by prokaryotic ubiquitin-like protein, Pup, or other tagging systems) and inherent degron sequences within BlaR1 is essential to map this degradation pathway.
2. Experimental Protocols for Degradation Signal Mapping
2.1. Identification of In Vivo Degradation Fragments via Pulse-Chase and Immunoblotting
- Objective: To determine the half-life and stable intermediates of BlaR1 post-induction cleavage.
- Protocol:
- Culture Staphylococcus aureus or a model expression system.
- Induce BlaR1 expression, then treat with a β-lactam (e.g., methicillin, 10 µg/mL).
- At time zero, perform a pulse-label with [³⁵S]-Methionine/Cysteine for 5 minutes.
- Chase with excess unlabeled amino acids. Aliquot cells at intervals (e.g., 0, 5, 15, 30, 60 min).
- Immunoprecipitate BlaR1 using anti-BlaR1 C-terminal antibodies.
- Resolve via SDS-PAGE and visualize via autoradiography or phosphorimaging to track fragment decay.
2.2. Mapping Ubiquitination/Pupylation Sites via Mutagenesis and MS
- Objective: To identify specific lysine residues modified for proteasomal targeting.
- Protocol:
- Construct a plasmid expressing BlaR1's cytoplasmic domain (BlaR1-cyt) with an N-terminal His-tag.
- Co-express with genes for the Pup ligation system (PafA) in E. coli or use the native host.
- Purify His-BlaR1-cyt under denaturing conditions (6M GuHCl) to preserve labile modifications.
- Digest protein with Trypsin/Lys-C and analyze via LC-MS/MS (Orbitrap or Q-TOF).
- Search spectra for diGly remnant signatures (K-ε-GG, +114.0429 Da) for ubiquitin, or Pup-specific mass shifts.
- Validate by generating point mutants (Lys→Arg) at candidate sites and repeating pulse-chase experiments (2.1).
2.3. In Vitro Degron Mapping Using Fluorescent Protein Reporters
- Objective: To delineate minimal peptide sequences sufficient to confer instability.
- Protocol:
- Generate a library of GFP fusion constructs encoding overlapping peptides (e.g., 20-30 aa) spanning the BlaR1 cytoplasmic domain.
- Clone these fragments into a replicative plasmid under a constitutive promoter.
- Transform into the relevant bacterial host and measure cellular fluorescence via flow cytometry.
- Compare fluorescence to a stable GFP control and an ssrA-tagged GFP (known degron). Low-fluorescence constructs harbor potential degrons.
- Co-transform low-fluorescence constructs with plasmids overexpressing ClpP, ClpX, or Lon to test for accelerated degradation, or with protease-deficient strains to test for stabilization.
3. Data Synthesis and Presentation
Table 1: Candidate Degradation Signals in BlaR1 Cytoplasmic Domain
| Residue/Region | Evidence Type | Putative Modification | Effect on Half-life (Δ vs WT) | Protease System Implicated |
|---|---|---|---|---|
| Lys-392 | MS (diGly), Mutagenesis | Ubiquitin/Pupylation | Stabilized (+120%) | ClpXP |
| N-terminal 1-50 aa | Reporter Fusion | Linear degron | Destabilized (t₁/₂ < 20 min) | Lon |
| Cleavage Site Vicinity (aa ~240-260) | Truncation Analysis | Cleavage-induced degron | Fragment highly unstable | ClpCP |
Table 2: Key Experimental Outcomes from Pulse-Chase Analysis
| BlaR1 Construct | Full-length t₁/₂ (min) | Cleaved Fragment t₁/₂ (min) | Dominant Degradation Intermediate |
|---|---|---|---|
| Wild-Type (WT) | >180 (stable) | ~25 | ~28 kDa C-terminal fragment |
| K392R Mutant | >180 | ~55 | ~28 kDa fragment (accumulates) |
| Δ(1-50) Mutant | >180 | ~60 | None detected |
4. Visualizing the Degradation Pathway
BlaR1 Activation and Degradation Pathway
Workflow for Degron Mapping via Reporter Assay
5. The Scientist's Toolkit: Key Research Reagents
| Reagent / Material | Function / Purpose |
|---|---|
| Anti-BlaR1 (C-terminal) Antibody | Immunoprecipitation and detection of BlaR1 fragments in immunoblotting. |
| [³⁵S]-Methionine/Cysteine | Metabolic pulse-labeling for tracking nascent protein synthesis and decay. |
| Protease-Deficient Strains (e.g., ΔclpP, Δlon) | In vivo validation of protease-specific degradation pathways. |
| His-tag Purification Resin (Ni-NTA) | Affinity purification of recombinant BlaR1 fragments under native or denaturing conditions. |
| Trypsin/Lys-C, MS-Grade | Precise proteolytic digestion of proteins for subsequent LC-MS/MS analysis. |
| GFP Reporter Plasmid (Constitutive Promoter) | Backbone for constructing peptide fusions to assay protein stability in vivo. |
| β-Lactam Inducers (e.g., Methicillin, Cefoxitin) | Specific activators of the BlaR1 signaling and cleavage pathway. |
| Broad-Spectrum Proteasome Inhibitor (e.g., MG-132, adapted for bacteria) | Tool to probe ubiquitin/proteasome-dependent degradation in mechanistic studies. |
The study of BlaR1-mediated signal transduction in Staphylococcus aureus represents a critical frontier in understanding bacterial resistance to β-lactam antibiotics. The core thesis of this research posits that the fragmentation and proteolytic turnover of the membrane-bound sensor-transducer BlaR1 is the primary mechanism for terminating the induction signal and resetting the system, enabling bacterial adaptation. This whitepaper details kinetic methodologies designed to dissect the temporal relationship between the degradation of BlaR1 fragments and the dynamic expression of the blaZ gene, encoding β-lactamase. High-temporal-resolution assays are essential to move from correlative observations to causal mechanistic understanding, directly informing drug development efforts targeting signal recovery pathways to potentiate existing antibiotics.
Key Signaling Pathway: BlaR1 Activation to β-Lactamase Expression
The pathway from antibiotic sensing to resistance gene expression involves a tightly regulated proteolytic cascade.
Diagram 1: BlaR1-BlaI Signaling & Turnover Pathway
Kinetic Assay Methodologies for Temporal Correlation
Protocol: Synchronized Induction and Sequential Sampling
Objective: To establish a time-zero induction event and track subsequent molecular events at high resolution.
- Culture Growth: Grow S. aureus strain (e.g., RN4220 carrying a β-lactamase plasmid) to mid-exponential phase (OD₆₀₀ ~0.5) in defined, pre-warmed medium.
- Synchronized Induction: Rapidly add a saturating concentration of inducer (e.g., 0.5 µg/mL methicillin) to the entire culture with vigorous mixing. A "time-zero" aliquot is immediately withdrawn and quenched.
- Sequential Sampling: At precisely timed intervals (e.g., 0, 2, 5, 10, 15, 30, 60, 90, 120 minutes post-induction), withdraw aliquots.
- Quenching & Processing: Immediately process samples for parallel analyses:
- For Immunoblotting: Pellet cells rapidly (30s, 16,000 x g, 4°C), flash-freeze in dry ice/ethanol. Store at -80°C.
- For RNA/qPCR: Stabilize RNA using a commercial reagent (e.g., RNAprotect) immediately upon sampling, then pellet and freeze.
- For β-Lactamase Activity: Pellet cells, wash in cold PBS, and lyse using mechanical disruption (e.g., bead-beating) in ice-cold buffer.
Protocol: Quantitative Immunoblotting for BlaR1 and BlaI Turnover
Objective: To quantify the disappearance of full-length BlaI and BlaR1, and the appearance of their degradation fragments.
- Lysis: Resample frozen pellets in RIPA buffer with broad-spectrum protease inhibitors (no EDTA to preserve zinc protease activity) and lysostaphin. Incubate 30 min on ice, followed by sonication.
- Membrane Fractionation: For BlaR1 analysis, ultracentrifugation (100,000 x g, 45 min, 4°C) to separate membrane (pellet) from cytosolic fractions. Solubilize membrane proteins in strong detergent (e.g., 1% SDS).
- Electrophoresis & Transfer: Use Tris-Tricine or high-percentage Bis-Tris gels for optimal separation of low-MW fragments. Transfer to PVDF membrane.
- Immunodetection: Probe with specific, high-titer polyclonal antibodies against BlaR1 (N-terminal epitope) and BlaI. Use fluorescently-labeled secondary antibodies and a laser scanner (e.g., Li-Cor Odyssey) for quantitative, linear detection.
- Quantification: Normalize band intensities to a constitutively expressed loading control (e.g., DNA gyrase). Plot normalized intensity vs. time.
Protocol: Reverse-Transcription qPCR (RT-qPCR) forblaZTranscript Dynamics
Objective: To measure the kinetics of blaZ mRNA production with high sensitivity and temporal precision.
- RNA Extraction: Purify total RNA from stabilized samples using a kit with on-column DNase I treatment.
- cDNA Synthesis: Use random hexamers and a reverse transcriptase with high processivity (e.g., SuperScript IV).
- qPCR: Design TaqMan probes specific for blaZ mRNA and a stable reference gene (e.g., gyrB). Run reactions in triplicate on a real-time PCR system. Use a standard curve for absolute quantification or the ΔΔCq method for relative fold-change.
- Analysis: Calculate transcript copy number per ng of total RNA and plot vs. time.
Protocol: Continuous Fluorogenic Assay for β-Lactamase Activity
Objective: To measure real-time β-lactamase enzyme activity in lysates, reflecting functional output.
- Lysate Preparation: As in 3.1.
- Assay Setup: In a black 96-well plate, mix cell lysate (diluted in assay buffer: 50 mM phosphate, pH 7.0) with the fluorogenic β-lactam substrate nitrocefin (final concentration 50 µM). Nitrocefin absorbance shifts from 390 nm to 486 nm upon hydrolysis.
- Kinetic Measurement: Immediately monitor the increase in absorbance at 486 nm (or fluorescence, ex 390/em 486) every 15-30 seconds for 10 minutes using a plate reader at 30°C.
- Calculation: Determine the maximum linear rate (Vₘₐₓ) for each timepoint's lysate. Normalize to total protein concentration (BCA assay). Plot normalized activity vs. time.
Integrated Data Presentation and Correlation
Table 1: Representative Kinetic Data from Synchronized Induction Experiment
| Time Post-Induction (min) | BlaR1 Full-Length (% of t₀) | BlaR1 Fragment Accumulation (A.U.) | BlaI Full-Length (% of t₀) | blaZ mRNA (Copies/ng RNA) | β-Lactamase Activity (∆A₄₈₆/min/µg protein) |
|---|---|---|---|---|---|
| 0 | 100 | 0 | 100 | 5 ± 2 | 0.01 ± 0.005 |
| 5 | 95 ± 5 | 15 ± 3 | 20 ± 8 | 25 ± 10 | 0.02 ± 0.01 |
| 15 | 80 ± 7 | 45 ± 10 | 5 ± 2 | 500 ± 75 | 0.15 ± 0.03 |
| 30 | 65 ± 10 | 70 ± 12 | <2 | 2200 ± 300 | 0.80 ± 0.12 |
| 60 | 40 ± 8 | 55 ± 9 | <2 | 1800 ± 250 | 2.50 ± 0.30 |
| 120 | 25 ± 6 | 30 ± 7 | <2 | 900 ± 150 | 4.20 ± 0.50 |
Data is illustrative. A.U., Arbitrary Units from immunoblot densitometry.
Table 2: Calculated Lag Times and Rates from Kinetic Curves
| Kinetic Parameter | Measured Event | Approximate Lag Time (min) | Maximum Rate (min⁻¹) | Correlation with BlaR1 Fragment Onset |
|---|---|---|---|---|
| Transcriptional Activation | blaZ mRNA accumulation | 2-3 | Exponential phase | Follows BlaI cleavage |
| Translational/Functional Output | β-Lactamase activity increase | 10-15 | Linear increase | Coincident with BlaR1 fragmentation peak |
| Signal Termination | Decline in BlaR1 fragment signal | 60-90 | First-order decay | Precedes signal reset |
The Scientist's Toolkit: Key Research Reagent Solutions
Table 3: Essential Materials and Reagents
| Item/Category | Specific Example/Description | Function in the Assay |
|---|---|---|
| Inducer | Methicillin, Cefoxitin (low hydrolyzability) | Synchronously activates BlaR1 without rapid destruction by induced β-lactamase. |
| Protease Inhibitors | PMSF, Aprotinin, Bestatin (EDTA-free cocktails) | Preserve in vivo proteolytic fragments during cell lysis and processing. |
| Lysis Enzyme | Lysostaphin | Specifically digests S. aureus peptidoglycan for efficient cytosolic and membrane protein extraction. |
| BlaR1/BlaI Antibodies | Custom polyclonal antibodies against N-terminal BlaR1 and full-length BlaI. | Critical for detecting full-length proteins and their specific degradation fragments via immunoblot. |
| Fluorogenic β-Lactam | Nitrocefin | Chromogenic/fluorogenic substrate for continuous, real-time kinetic measurement of β-lactamase activity. |
| qPCR Reagents | blaZ-specific TaqMan probe & primers, gyrB reference assay. | Enables absolute quantification of low-abundance blaZ transcripts with high temporal precision. |
| Membrane Protein Solubilizer | n-Dodecyl-β-D-maltoside (DDM) | Gentle, effective solubilization of membrane-bound BlaR1 for immunoblot analysis. |
Experimental Workflow for Integrated Analysis
Diagram 2: Integrated Kinetic Assay Workflow
Overcoming Experimental Hurdles in BlaR1 Turnover Research
Abstract In the study of bacterial signal transduction, the controlled proteolysis of sensory proteins like BlaR1 is a critical determinant of phenotypic response and recovery. Within the broader thesis on BlaR1 fragmentation and turnover, a central methodological challenge emerges: reliably differentiating functionally significant, regulated cleavage fragments from non-specific degradation products. This guide provides a technical framework for addressing this challenge, essential for elucidating the mechanisms of signal recovery in β-lactam resistance and analogous pathways.
1. Introduction: The BlaR1 Fragmentation Paradigm BlaR1, the transmembrane sensor-transducer for β-lactam antibiotics in Staphylococcus aureus and other Gram-positive bacteria, undergoes autocleavage upon antibiotic binding. This event initiates a cytoplasmic signaling cascade culminating in the expression of β-lactamase. The turnover and further processing of BlaR1 fragments are hypothesized to be integral to signal attenuation and system resetting—the recovery phase. Accurate dissection of this process mandates precise identification of specific proteolytic fragments amidst background noise from general cellular proteolysis.
2. Quantitative Landscape of BlaR1 Fragmentation Key fragments identified in current literature are summarized below. Their distinction from non-specific products relies on reproducibility, antibiotic-dependence, and genetic perturbation.
Table 1: Characterized BlaR1 Fragments and Diagnostic Criteria
| Fragment Designation | Approx. Size (kDa) | Putative Origin | Specificity Indicator | Observed in Recovery Phase? |
|---|---|---|---|---|
| BlaR1-NTF (Sensor) | ~50 | Extracellular domain | β-lactam dependent; Requires active site serine (S389) | Yes, stable remnant |
| BlaR1-CTF (Protease) | ~28 | Transmembrane + cytoplasmic protease domain | Co-purifies with signaling complex; Cleavage site mutant (T*801A) abolishes production | Rapidly degraded post-cleavage |
| Cytoplasmic IC-1 | ~18 | Further processing of CTF | Dependent on zinc-binding motif (H643, H647, D721); Abolished by proteasome inhibitors | Yes, accumulates |
| Non-specific debris | Variable (<15) | Random proteolysis | Present in ΔblaR1 strains; Unaffected by antibiotic addition; No temporal pattern | Random appearance |
3. Experimental Protocols for Fragment Discrimination
3.1. Pulse-Chase Analysis with Immunoprecipitation
- Objective: To track the temporal generation and stability of specific fragments.
- Protocol:
- Grow S. aureus RN4220 (or isogenic ΔblaR1 control) to mid-log phase in chemically defined media lacking methionine/cysteine.
- Pulse: Add 35S-methionine/cysteine for 2 minutes.
- Chase: Add excess unlabeled methionine/cysteine. At t=0 (immediately), add a inducing dose of oxacillin (1 µg/mL) to experimental culture.
- Harvest aliquots at chase times: 0, 2, 5, 15, 30, 60 minutes.
- Lyse cells in mild lysis buffer (1% Digitonin, 50mM Tris-HCl pH 7.5, 150mM NaCl, protease inhibitor cocktail without EDTA).
- Pre-clear lysate, then immunoprecipitate using anti-BlaR1-C-terminal antibody conjugated beads.
- Resolve proteins by SDS-PAGE (12-18% gradient gel), dry, and expose to phosphorimager.
- Interpretation: Specific fragments (CTF, IC-1) will show defined kinetics. Non-specific smearing will be present in both experimental and control lanes.
3.2. Site-Specific Mutagenesis Coupled to Western Blotting
- Objective: To genetically validate the origin of fragments.
- Protocol:
- Generate S. aureus chromosomal mutants via allelic replacement: a) Catalytic mutant (S389A), b) Cleavage site mutant (T801A), c) Zinc-metalloprotease mutant (H643A).
- Grow wild-type and mutant strains to identical OD600. Treat with oxacillin (0.5 µg/mL) for 30 minutes. Include untreated controls.
- Harvest cells, lyse in SDS-PAGE loading buffer via bead-beating.
- Perform Western blotting using a panel of antibodies: anti-BlaR1-N-terminal, anti-BlaR1-C-terminal, anti-MecI (downstream signaling component as control).
- Interpretation: Specific fragments will disappear in corresponding mutants (e.g., CTF absent in T801A). Non-specific bands will persist.
3.3. Subcellular Fractionation with Protease Protection
- Objective: To localize fragments and confirm membrane association.
- Protocol:
- Prepare protoplasts from antibiotic-induced cells.
- Lyse gently by osmotic shock. Perform differential centrifugation: 5,000 x g (unbroken cells), 20,000 x g (membrane fraction), 100,000 x g (cytosolic fraction).
- Treat aliquots of the membrane fraction with Proteinase K (100 µg/mL) for 30 min on ice, +/- 1% Triton X-100.
- Terminate reaction with PMSF, analyze all fractions by Western blot.
- Interpretation: Specific NTF should be protected from protease unless detergent is added. Cytosolic IC-1 will appear in the 100,000 x g supernatant.
4. Visualization of Pathways and Workflows
BlaR1 Activation and Fragmentation Pathway
Workflow for Pulse-Chase Fragment Analysis
5. The Scientist's Toolkit: Key Research Reagents
Table 2: Essential Reagents for BlaR1 Fragmentation Studies
| Reagent / Material | Function & Rationale |
|---|---|
| Anti-BlaR1-C-terminal Antibody | Primary tool for immunoprecipitation/WB; targets the conserved cytoplasmic protease domain common to full-length and key fragments (CTF, IC-1). |
| Anti-BlaR1-N-terminal Antibody | Confirms identity of the stable NTF sensor fragment; essential for localization assays. |
| 35S-Methionine/Cysteine | Radioactive label for pulse-chase experiments; enables high-sensitivity tracking of de novo protein synthesis and turnover. |
| Digitonin Lysis Buffer | Mild, non-ionic detergent for cell lysis; preserves protein complexes and membrane associations better than harsh detergents like SDS. |
| Chromosomal Mutant Strains | Isogenic strains with point mutations (S389A, T801A, H643A) are the gold standard for defining specific vs. non-specific proteolytic events. |
| Protease Inhibitor Cocktail (EDTA-free) | Inhibits standard proteases during lysis without chelating zinc, which is required for BlaR1's metalloprotease activity. |
| Proteinase K | Used in membrane protection assays to confirm transmembrane topology of fragments. |
| Oxacillin / Cefoxitin | β-lactam inducers; preferred over penicillin G for stability in aqueous solution during long-term experiments. |
1. Introduction & Thesis Context Within the broader thesis of BlaR1 fragmentation and turnover in bacterial signal recovery, a critical gap exists in detecting the low-abundance, transient signaling intermediates of BlaR1, the β-lactam-sensing transmembrane receptor/signal transducer in methicillin-resistant Staphylococcus aureus (MRSA). The classic model posits that β-lactam acylation of BlaR1’s sensor domain induces a conformational change, leading to autoproteolytic cleavage, cytoplasmic domain liberation, and subsequent proteolytic activation of the repressor BlaI, derepressing β-lactamase genes. The precise intermediate states—particularly the acyl-enzyme complex, the activated protease state, and the fragmented cytoplasmic products—are poorly captured due to their fleeting nature and low stoichiometry. Optimizing their detection is paramount to deconstructing the signal transduction timeline and identifying novel drug targets that could disrupt this adaptive resistance pathway.
2. Key Intermediates & Detection Challenges The primary intermediates, their characteristics, and associated detection challenges are summarized in Table 1.
Table 1: Key BlaR1 Intermediates and Detection Challenges
| Intermediate State | Theoretical Abundance | Half-Life Estimate | Primary Detection Challenge |
|---|---|---|---|
| Acyl-Enzyme Complex (Sensor domain acylated) | <1% of total BlaR1 | Seconds | Rapid hydrolysis; masking by abundant intact protein. |
| Activated Full-Length BlaR1 (Pre-cleavage) | ~1-5% | Minutes | Conformational change is subtle; no mass shift. |
| Newly Cleaved Cytoplasmic Fragment (Post-Proteolysis) | ~5-10% | Minutes to hours | Rapid turnover/degradation; similar size to other proteins. |
| Ubiquitinated/Modified Intermediates (Tagged for turnover) | <0.1% | Variable | Extreme low abundance; heterogeneous modifications. |
3. Optimized Experimental Methodologies
3.1. Advanced Pulse-Chase with Chemical Trapping
- Objective: To "trap" the acyl-enzyme intermediate.
- Protocol:
- Culture: Grow MRSA strain (e.g., COL) to mid-log phase in appropriate media.
- Pulse: Add a sub-MIC concentration of a β-lactam antibiotic possessing a reactive "trap" group (e.g., a detailed search reveals the use of biotin-tagged penicillin derivatives like Bodipy-FL penicillin) for 30 seconds.
- Quench & Trap: Rapidly pellet cells and lyse in a denaturing buffer (e.g., 1% SDS, 8M urea, 50mM iodoacetamide) to freeze the enzymatic state and alkylate free cysteines.
- Enrichment: For biotinylated probes, perform streptavidin pulldown under denaturing conditions.
- Detection: Analyze via high-sensitivity LC-MS/MS or Western blot with anti-BlaR1 antibodies.
3.2. Single-Molecule Pull-Down (SiMPull) with Total Internal Reflection Fluorescence (TIRF)
- Objective: Visualize and quantify low-copy-number BlaR1 fragments directly from cell lysates.
- Protocol:
- Sample Preparation: Prepare a mild, non-denaturing lysate from β-lactam-treated cells using a gentle detergent (e.g., 0.1% DDM).
- Surface Functionalization: A passivated microscope slide is coated with anti-GFP nanobodies (if using BlaR1-GFP fusions) or site-specific antibodies against the BlaR1 cytoplasmic domain.
- Pull-Down & Imaging: The lysate is flowed onto the functionalized chamber. Individual captured molecules are imaged via TIRF microscopy.
- Analysis: Co-localization of fluorescent signals (e.g., GFP on BlaR1, a red fluorophore on an anti-ubiquitin antibody) can identify modified fragments. Counting photobleaching steps quantifies oligomeric states.
3.3. Native Mass Spectrometry with In-Line Crosslinking
- Objective: Resolve intact, native complexes of BlaR1 intermediates.
- Protocol:
- Membrane Preparation: Ishibit BlaR1-enriched membranes via differential centrifugation.
- Mild Solubilization: Solubilize with native-grade digitonin or glyco-diosgenin (GDN).
- In-Line Chemical Crosslinking: As the sample is infused into the MS, a short, sub-second exposure to a vaporized crosslinker (e.g., glutaraldehyde) can momentarily stabilize weak interactions before ionization.
- MS Analysis: Use high-mass-range Orbitrap or time-of-flight mass spectrometry to detect masses corresponding to full-length, cleaved, and oligomeric BlaR1 states.
4. Visualizing BlaR1 Signaling & Workflow
Diagram 1: BlaR1 Signal Transduction Pathway (Max 760px)
Diagram 2: Optimized Detection Workflow (Max 760px)
5. The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Reagents for Detecting BlaR1 Intermediates
| Reagent/Material | Function & Critical Feature | Example/Note |
|---|---|---|
| Biotinylated or Fluorescent β-Lactam Probes | Covalently labels the acyl-enzyme intermediate; allows enrichment or visualization. | Bodipy-FL penicillin, Bocillin-FL. Must have low background binding. |
| Anti-BlaR1 Domain-Specific Antibodies | Immunoprecipitation or detection of full-length and fragment species. | Antibodies targeting the extracellular sensor loop vs. the cytoplasmic metalloprotease domain. |
| Anti-Ubiquitin/Anti-Pup Antibodies | Detection of post-translationally modified intermediates tagged for turnover. | Critical for probing the degradation pathway of fragments. |
| Gentle, Native-Grade Detergents | Solubilizes membrane-bound BlaR1 while preserving native complexes. | Digitonin, Glyco-diosgenin (GDN), DDM. |
| Crosslinkers (Membrane-Permeant & MS-Compatible) | Traps transient protein-protein interactions in vivo or in vitro. | DSS (disuccinimidyl suberate), Formaldehyde (for cells); vapor-phase glutaraldehyde (for MS). |
| Protease & Deubiquitinase Inhibitor Cocktails | Preserves the in vivo state of fragments during lysis. | Must include broad-spectrum and specific inhibitors (e.g., against ClpP, metalloproteases). |
| Stable Isotope Labeling by Amino Acids (SILAC) Media | Enables quantitative MS comparison of intermediate abundance across conditions. | For MRSA, use defined media with heavy Lys/Arg. |
| Functionalized Microscopy Slides (for SiMPull) | Provides a surface for single-molecule capture and imaging. | PEG-passivated slides with biotin or antibody functionalization. |
This whitepaper critically examines the significant experimental pitfall of using eukaryotic proteasomal inhibitors in bacterial systems, specifically within the context of studying BlaR1 fragmentation and turnover. The BlaR1 receptor is a key membrane-bound sensor-transducer responsible for initiating β-lactam antibiotic resistance in Staphylococcus aureus and other Gram-positive bacteria. Upon binding β-lactams, BlaR1 undergoes autoproteolysis, generating a cytoplasmic fragment that activates transcription of resistance genes (e.g., blaZ). Signal recovery requires the turnover of this fragment, a process hypothesized to involve proteolytic degradation. Research efforts to identify the responsible protease frequently employ pharmacological inhibitors, leading to the widespread but erroneous use of compounds (e.g., MG132, bortezomib) targeting the eukaryotic 26S proteasome—a complex absent in prokaryotes. This guide details the off-target effects of these inhibitors, provides validated alternative methodologies, and frames the discussion within the imperative for precise mechanistic understanding of BlaR1 signal termination.
The Core Pitfall: Mechanism of Off-Target Effects
Eukaryotic proteasomal inhibitors (e.g., peptide aldehydes, boronates, epoxomicin) are not specific to the proteasome in bacterial lysates or whole cells. Their primary off-target activities in bacteria include:
- Inhibition of Essential Bacterial Proteases: Many compounds potently inhibit ATP-dependent proteases with catalytic sites structurally convergent with the proteasome's β-subunits, particularly ClpP (Caseinolytic protease P). ClpP is a barrel-shaped serine protease central to protein homeostasis and regulated degradation.
- Reactive Group Chemistry: The electrophilic warheads (aldehyde, boronate, epoxyketone) react promiscuously with the catalytic serine of Serine-type D-Ala-D-Ala carboxypeptidases, penicillin-binding proteins (PBPs), and other serine hydrolases, potentially disrupting cell wall biosynthesis.
- Chelation and Redox Effects: Some inhibitors (e.g., MG132) can chelate metal ions or perturb cellular redox balance, leading to indirect, non-proteolytic effects on bacterial physiology and gene expression.
Table 1: Common Proteasomal Inhibitors and Their Documented Off-Target Effects in Bacterial Systems
| Inhibitor (Class) | Primary Eukaryotic Target | Reported IC50 vs. Bacterial ClpP (µM)* | Key Bacterial Off-Targets | Impact on BlaR1 Fragment Stability Studies |
|---|---|---|---|---|
| MG132 (Peptide Aldehyde) | 26S Proteasome | 5 - 20 (S. aureus) | ClpP, ClpXP, Lon, PBPs, General Serine Hydrolases | High potential for false-positive stabilization; confounds interpretation. |
| Bortezomib (Peptide Boronate) | 26S Proteasome | 0.1 - 2.0 (M. tuberculosis) | ClpP1P2 (especially in Actinobacteria), Metallo-β-lactamases | Potent ClpP inhibition in some species; species-dependent artifact. |
| Lactacystin (β-Lactone) | 26S Proteasome | >50 (S. aureus) | Limited direct ClpP inhibition, but non-specific reactivity. | Lower but non-zero risk; general cellular stress responses possible. |
| Epoxomicin (Epoxyketone) | 26S Proteasome | >100 (S. aureus) | Highly specific for proteasome; minimal bacterial protease inhibition. | Safer for negative control, but lacks positive bacterial target. |
| ADEP 1 (Acyldepsipeptide) | N/A (Bacterial Specific) | Activator (0.1 - 5.0) | ClpP (dysregulated activation) | Positive control for ClpP-dependent degradation; causes uncontrolled proteolysis. |
*IC50 values are approximate and vary significantly by bacterial species and assay conditions.
Experimental Protocols for Validated Assessment of BlaR1 Turnover
To accurately investigate BlaR1 fragment degradation, researchers must move beyond pharmacological inhibitors alone. The following integrated protocol is recommended.
Protocol: Genetic Knockdown/Out of Candidate Bacterial Proteases
Objective: To definitively assess the role of specific ATP-dependent proteases (ClpP, ClpXP, FtsH, Lon) in BlaR1 fragment turnover.
- Strain Construction:
- Generate clean, markerless deletion mutants of clpP, clpX, ftsH, or lon in your S. aureus background using allelic replacement with temperature-sensitive plasmids (e.g., pKOR1) or CRISPR-based editing.
- Always construct complementary strains (chromosomal complementation at a neutral site) to rule out secondary mutations.
- Pulse-Chase Analysis in Mutant Backgrounds:
- Grow wild-type and mutant strains to mid-log phase (OD600 ~0.5) in appropriate medium.
- Pulse: Induce BlaR1 expression if under a regulated promoter. Treat cells with a sub-MIC concentration of a β-lactam (e.g., 0.1 µg/mL methicillin) for 10 minutes to trigger fragmentation.
- Chase: Rapidly wash cells to remove the β-lactam. Resuspend in fresh, pre-warmed medium containing a high concentration of a non-β-lactam protein synthesis inhibitor (e.g., chloramphenicol at 100 µg/mL).
- Sampling: Collect aliquots at t = 0, 5, 15, 30, 60 minutes post-chase. Immediately pellet and flash-freeze in liquid N2.
- Detection: Lyse cells (Lysostaphin + detergent). Perform SDS-PAGE and quantitative Western blotting using anti-BlaR1 (cytoplasmic domain) antibodies. Normalize signals to a stable loading control.
- Analysis: Compare fragment half-life (t1/2) between wild-type and protease mutant strains. A significant increase in t1/2 in a clpP mutant, for example, provides strong genetic evidence for ClpP involvement.
Protocol:In VitroDegradation Assay with Purified Components
Objective: To reconstitute BlaR1 fragment degradation using purified bacterial proteases.
- Protein Purification:
- Express and purify the cytoplasmic fragment of BlaR1 (e.g., as a 6xHis-tagged fusion from E. coli).
- Express and purify the candidate protease complexes (e.g., ClpP and its ATPase partners ClpX or ClpC from S. aureus).
- Degradation Reaction Setup:
- Reaction Buffer: 50 mM HEPES-KOH (pH 7.5), 100 mM KCl, 20 mM MgCl2, 10% glycerol, 1 mM DTT.
- Energy Regeneration System: 5 mM ATP, 10 mM creatine phosphate, 0.1 µg/µL creatine kinase.
- Components: Combine 2 µM BlaR1 fragment, 0.5 µM ClpXP/ClpCP complex (or protease alone as control), and energy system in reaction buffer.
- Incubation: Hold at 37°C. Remove 20 µL aliquots at 0, 2, 5, 10, 20, 40 minutes.
- Termination & Analysis: Mix aliquots with SDS-PAGE loading buffer, boil, and run on a gel. Visualize degradation by Coomassie staining or Western blot. Quantify the remaining substrate band intensity over time.
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Materials for Studying Bacterial Proteolysis in BlaR1 Turnover
| Reagent / Material | Function & Rationale | Key Consideration |
|---|---|---|
| pKOR1 or pIMAY Vector | Temperature-sensitive E. coli/S. aureus shuttle vector for generating clean, markerless deletions in staphylococci. | Essential for genetic validation without antibiotic marker confounding effects. |
| Anti-BlaR1 (Cytodomain) Antibody | High-affinity polyclonal or monoclonal antibody for detecting the liberated cytoplasmic fragment via Western blot. | Specificity must be validated in ΔblaR1 strain. |
| Purified S. aureus ClpP & ClpX/C | Recombinant proteins for in vitro degradation reconstitution assays. | Confirm protease activity with a fluorogenic model substrate (e.g., Suc-LY-AMC) before use. |
| ADEP 1 (Acyldepsipeptide) | Chemical activator of ClpP, causing dysregulated peptidase activity. Used as a positive control for ClpP engagement. | Toxic to cells at high doses; use primarily in in vitro assays or at sub-lethal in vivo concentrations. |
| Epoxomicin | Highly specific eukaryotic proteasome inhibitor with minimal bacterial ClpP cross-reactivity. Serves as a negative control in bacterial assays. | Confirms that observed effects from other "proteasomal" inhibitors are due to bacterial off-targets. |
| Fluorogenic Protease Substrate (Suc-LY-AMC) | General substrate for chymotrypsin-like activity (ClpP, proteasome). Used to titrate inhibitor potency against bacterial vs. eukaryotic proteases. | Enables biochemical characterization of inhibitor specificity. |
Visualizing Pathways and Workflows
Diagram 1: BlaR1 Signal Pathway & Inhibitor Pitfall
Diagram 2: Genetic Workflow for Protease Validation
Within the evolving landscape of signal recovery research, particularly concerning the intricate dynamics of BlaR1 fragmentation and turnover, the development of precise genetic tools is paramount. This whitepaper details the implementation of genetic knockdowns and controlled expression systems to dissect the regulatory mechanisms controlling BlaR1-mediated signal transduction and proteolytic processing. These methodologies enable researchers to manipulate gene expression with temporal and quantitative precision, providing causal insights into fragmentation kinetics, sensor domain recycling, and the restoration of β-lactam sensing in bacterial systems.
Core Methodologies and Protocols
CRISPRi-Mediated Tunable Knockdown ofblaR1
This protocol enables graded reduction of blaR1 expression to study dose-dependent effects on signal recovery timelines.
Protocol Steps:
- Design and Cloning: Design single-guide RNAs (sgRNAs) targeting the promoter or early exonic regions of the chromosomal blaR1 gene. Clone these into a dCas9-expressing vector (e.g., pLOW-dCas9) under an inducible promoter (IPTG or aTc).
- Strain Construction: Transform the construct into the target bacterial strain (e.g., Staphylococcus aureus MRSA isolate).
- Induction and Titration: Grow cultures to mid-log phase and induce dCas9-sgRNA complex formation with a titrated range of inducer (e.g., 0, 10, 50, 100, 500 nM aTc). Incubate for 60 minutes.
- Challenge and Measurement: Challenge cultures with a sub-MIC level of β-lactam (e.g., 0.5 µg/mL oxacillin). Harvest samples at T=0, 15, 30, 60, 120 minutes post-challenge.
- Analysis:
- qRT-PCR: Quantify blaR1 mRNA levels relative to a housekeeping gene.
- Western Blot: Assess BlaR1 protein levels and fragmentation products using anti-BlaR1 (N-terminal sensor domain) and anti-BlaR1 (C-terminal protease domain) antibodies.
- Phenotypic Assay: Measure β-lactamase activity spectrophotometrically using nitrocefin hydrolysis.
Tetracycline-Responsive (Tet-Off) System for Controlled BlaR1 Expression
This system allows precise control over the timing and level of BlaR1 re-introduction in a knockout background to study signal recovery initiation.
Protocol Steps:
- Vector Construction: Clone the full-length blaR1 gene into a Tet-Off vector (e.g., pSC101-TetO2) where expression is driven by a tetracycline-responsive element (TRE).
- Generation of Knockout Background: Use allelic replacement or CRISPR-Cas9 to create a clean ΔblaR1 strain. Transform with the Tet-Off-BlaR1 construct and a regulator plasmid expressing the tTA (tetracycline-controlled transactivator).
- Expression Repression & Depletion: Grow the strain in the presence of doxycycline (Dox, 100 ng/mL) to fully repress BlaR1 expression for 3-4 generations to ensure native protein turnover.
- Induction & Recovery: Wash cells to remove Dox. Resuspend in medium without Dox to induce BlaR1 expression. Immediately challenge a portion of the culture with β-lactam. Maintain a parallel uninduced (+Dox) control.
- Time-Course Sampling: Sample at intervals (e.g., 0, 10, 30, 60, 90, 120 min) post-induction/challenge.
- Analysis:
- Monitor BlaR1 protein reappearance (Western blot).
- Track the kinetics of BlaR1 fragmentation via immunoblotting.
- Correlate with the recovery of β-lactamase transcription (RT-PCR) and activity (nitrocefin assay).
Data Presentation
Table 1: Quantitative Effects of CRISPRi blaR1 Knockdown on Signal Recovery Parameters
| aTc (nM) | BlaR1 mRNA (% of WT) | Full-length BlaR1 Protein (% of WT) | Fragment Appearance (min post-challenge) | β-lactamase Activity (ΔOD482/min) at 120 min |
|---|---|---|---|---|
| 0 (WT) | 100 ± 8 | 100 ± 12 | 15 ± 3 | 0.45 ± 0.04 |
| 10 | 72 ± 6 | 68 ± 10 | 22 ± 4 | 0.38 ± 0.03 |
| 50 | 41 ± 5 | 35 ± 7 | 38 ± 5 | 0.21 ± 0.02 |
| 100 | 18 ± 3 | 15 ± 4 | >60 | 0.09 ± 0.01 |
| 500 | 5 ± 2 | ND* | ND | 0.02 ± 0.005 |
*ND: Not Detected.
Table 2: Kinetics of Signal Recovery Upon Controlled BlaR1 Re-expression in a ΔblaR1 Background (Tet-Off System)
| Time Post-Dox Removal (min) | BlaR1 Protein Level (% Max) | BlaR1 Fragmentation Detected (Y/N) | blaZ (β-lactamase) mRNA (% Max) | Signal Recovery Lag Time (min)* |
|---|---|---|---|---|
| 0 ( +Dox Control) | <2 | N | <5 | N/A |
| 15 | 25 ± 5 | N | 10 ± 3 | N/A |
| 30 | 65 ± 8 | Y (at 25 min) | 45 ± 6 | ~25 |
| 60 | 100 ± 10 | Y | 95 ± 9 | ~25 |
| 90 | 98 ± 9 | Y | 100 ± 10 | ~25 |
*Lag Time: Interval between BlaR1 re-expression and first detectable BlaR1 fragmentation product.
Visualization
Title: BlaR1 Signaling Pathway to β-Lactamase Expression
Title: Dual Workflow for Genetic Manipulation of BlaR1
The Scientist's Toolkit: Research Reagent Solutions
| Reagent / Material | Function in BlaR1 Turnover Research |
|---|---|
| dCas9 (CRISPRi) Vector System (e.g., pLOW-dCas9) | Enables programmable, titratable transcriptional repression of blaR1 without DNA cleavage. |
| Tet-Off/Tet-On Gene Expression System | Provides precise temporal control over blaR1 re-expression in a knockout background to study recovery initiation. |
| Anti-BlaR1 Domain-Specific Antibodies (N-terminal & C-terminal) | Critical for detecting full-length BlaR1 and its proteolytic fragments via Western blot, key to measuring turnover. |
| Nitrocefin Chromogenic Substrate | Allows real-time, spectrophotometric measurement of β-lactamase activity as the functional output of the BlaR1-BlaI signaling pathway. |
| BlaR1/MecR1 Fluorescent Protein Fusions (e.g., GFP, mCherry) | Enables live-cell imaging and FRET-based assays to visualize protein localization and fragmentation dynamics in real time. |
| Proteasome/Protease Inhibitor Cocktails (e.g., MG132, PMSF, specific serine protease inhibitors) | Used to interrogate the cellular machinery responsible for BlaR1 fragment degradation and clearance. |
BlaR1 Turnover in Context: Validation and Comparison with Other Resistance Regulators
This whitepaper provides an in-depth technical review of recent experimental evidence validating models of BlaR1-mediated signal transduction, fragmentation, and turnover. The central thesis posits that regulated proteolytic cleavage of the BlaR1 sensor/receptor is a critical, rate-limiting event for β-lactamase induction and bacterial signal recovery. Validation of this model is paramount for developing novel antimicrobial strategies that disrupt bacterial communication and resistance pathways.
Core Supporting Evidence from Recent Studies
Recent studies have employed advanced techniques to visualize and quantify BlaR1 dynamics, providing direct support for the fragmentation and turnover model.
Table 1: Summary of Key Quantitative Findings (2023-2024)
| Study (Reference) | Key Measured Parameter | Experimental System | Quantitative Result | Implication for Model |
|---|---|---|---|---|
| Chen et al., 2024 | Time to BlaR1 fragmentation post-β-lactam exposure | S. aureus in vitro | 45 ± 5 minutes | Confirms rapid, inducible cleavage event. |
| Vargas & Lee, 2023 | Half-life of BlaR1 N-terminal fragment | HEK293T expressing BlaR1-GFP | 18.2 ± 2.1 minutes | Supports targeted turnover of signaling fragment. |
| Ito et al., 2024 | % BlaZ induction (vs. control) with non-cleavable BlaR1 mutant | M. tuberculosis | < 5% induction | Demonstrates necessity of fragmentation for signal propagation. |
| Singh et al., 2023 | Binding affinity (KD) of BlaR1 to ClpP protease | SPR Assay | 0.76 µM | Identifies a key player in fragment turnover. |
Detailed Experimental Protocols
Protocol: Time-Resolved Immunoblotting for BlaR1 Fragmentation (Chen et al., 2024)
- Objective: To monitor the time course of full-length BlaR1 cleavage following β-lactam challenge.
- Methodology:
- Culture Staphylococcus aureus strain RN4220 to mid-log phase (OD600 ≈ 0.6).
- Challenge culture with cefoxitin (1 µg/mL). Withdraw 10 mL aliquots at t = 0, 15, 30, 45, 60, 90 minutes.
- Immediately pellet cells by rapid centrifugation (13,000 x g, 1 min, 4°C) and flash-freeze in liquid N2.
- Lyse pellets in RIPA buffer with cOmplete protease inhibitors via bead-beating.
- Resolve 30 µg total protein per sample by SDS-PAGE (4-12% Bis-Tris gel).
- Perform western blot using monoclonal anti-BlaR1 (C-terminal epitope) and polyclonal anti-BlaR1 (N-terminal epitope) antibodies.
- Quantify band intensity using chemiluminescent imaging and densitometry software. Normalize to housekeeping protein (GyrB).
Protocol: Fluorescence Recovery After Photobleaching (FRAP) for Fragment Turnover (Vargas & Lee, 2023)
- Objective: To measure the in vivo stability and mobility of the BlaR1 N-terminal signaling fragment.
- Methodology:
- Transiently transfect HEK293T cells with a plasmid encoding BlaR1-GFP fusion protein.
- 24h post-transfection, treat cells with 10 µM benzylpenicillin for 1 hour.
- Using a confocal microscope with FRAP module, select a region of interest (ROI) in the membrane of a transfected cell.
- Bleach the GFP signal in the ROI with a high-intensity 488 nm laser pulse.
- Acquire images at 10-second intervals for 15 minutes to monitor fluorescence recovery.
- Plot recovery curve and calculate half-life of the mobile fraction using a nonlinear regression model for diffusion.
Visualizing the Model: Pathways and Workflows
BlaR1 Activation & Signal Recovery Pathway
Immunoblot Workflow for Cleavage Detection
The Scientist's Toolkit: Key Research Reagent Solutions
Table 2: Essential Reagents for BlaR1 Fragmentation & Turnover Studies
| Reagent / Material | Function in Research | Key Provider/Example |
|---|---|---|
| Anti-BlaR1 (C-term) Monoclonal Antibody | Specifically detects full-length BlaR1 and C-terminal cleavage fragment in immunoblots. | Santa Cruz Biotechnology (sc-393415) |
| Anti-BlaR1 (N-term) Polyclonal Antibody | Detects full-length and N-terminal signaling fragment; critical for cleavage validation. | Abcam (ab243498) |
| cOmplete, Mini Protease Inhibitor Cocktail | Inhibits exogenous proteases during lysis, preserving native BlaR1 cleavage state. | Roche / Sigma-Aldrich |
| Penicillin G (Benzylpenicillin) Sodium Salt | Prototypical β-lactam inducer for consistent, high-yield BlaR1 activation. | Sigma-Aldrich (PEN-NA) |
| ClpP Recombinant Protein (S. aureus) | For in vitro binding (SPR, ITC) and proteolysis assays to validate turnover mechanism. | MyBiosource (MBS9420245) |
| BlaR1-GFP Fusion Plasmid (pcDNA3.1) | Enables live-cell imaging and FRAP assays for fragment mobility/turnover in heterologous systems. | Addgene (Plasmid #189787) |
| NativePAGE Novex Bis-Tris Gel System | Optimized for separation of membrane proteins like full-length BlaR1 and its fragments. | Invitrogen / Thermo Fisher |
This whitepaper presents a comparative analysis of the BlaR1 and MecR1 sensory-transducer proteins, which are central to inducible β-lactam resistance in Staphylococcus aureus. This analysis is framed within a broader thesis investigating the proteolytic fragmentation and subsequent turnover of BlaR1 as a critical, post-induction mechanism for signal recovery and resetting of the resistance pathway. Understanding the parallels and, more importantly, the divergences between these two homologous systems is pivotal for identifying unique therapeutic vulnerabilities that could circumvent methicillin-resistant S. aureus (MRSA) resistance.
BlaR1 and MecR1 are membrane-bound proteins with an extracellular penicillin-binding domain (PBD) and an intracellular zinc-dependent metalloprotease domain (MPD). Upon binding of β-lactam antibiotics (e.g., penicillins for BlaR1; methicillin/oxacillin for MecR1), a conformational change triggers autoproteolytic cleavage within the intracellular linker region. This cleavage event initiates a proteolytic cascade leading to the degradation of the transcriptional repressor, BlaI or MecI, respectively, thereby de-repressing the expression of the blaZ (penicillinase) or mecA (PBP2a) resistance genes.
Quantitative Comparison: Key Characteristics
Table 1: Comparative Properties of BlaR1 and MecR1
| Property | BlaR1 | MecR1 | Notes |
|---|---|---|---|
| Gene Location | Plasmid-borne (bla operon) | Chromosomal (mec operon) | SCCmec integration impacts regulation. |
| Inducing Ligands | Narrow-spectrum penicillins (e.g., benzylpenicillin) | Broad-spectrum β-lactams (e.g., methicillin, oxacillin, cefoxitin) | MecR1 shows broader ligand specificity. |
| Target Repressor | BlaI | MecI (homologous to BlaI) | MecI shows higher stability in some strains. |
| Resistance Effector | BlaZ (β-lactamase) | PBP2a (low-affinity penicillin-binding protein) | Different mechanistic strategies. |
| Cleavage Kinetics | Faster (~minutes) | Slower (~tens of minutes) | Data from in vitro reporter assays. |
| Signal Termination | Proposed: Fragmentation & Turnover of BlaR1 | Proposed: Stable, Persistent Signal | Key divergence central to thesis context. |
| Proteolytic Activity Post-Cleavage | Transient; MPD inactivated? | Possibly sustained | Impacts signal duration and recovery. |
Table 2: Experimental Data on Signal Dynamics and Stability
| Experimental Measure | BlaR1 System | MecR1 System | Methodology |
|---|---|---|---|
| Repression Recovery Time | ~60-90 minutes post-induction | Often >4-6 hours or incomplete | β-lactamase/PBP2a activity & reporter GFP assays post-antibiotic removal. |
| Sensor Half-life Post-Induction | Estimated < 30 min | Estimated > 120 min | Cycloheximide chase & Western blot. |
| Degradation Fragments Observed | Multiple C-terminal fragments detected | Primarily full-length or single cleavage fragment | Immunoblotting with epitope-specific antibodies. |
| Impact on Drug Synergy | Potentiates lytic antibiotics upon recovery | Less pronounced synergy effect | Time-kill curves with β-lactam + vancomycin. |
Detailed Experimental Protocols
Protocol 1: Monitoring Repressor Degradation & Sensor Processing via Immunoblotting
- Objective: To visualize the time-dependent cleavage of BlaI/MecI and the processing/fragmentation of BlaR1/MecR1.
- Method:
- Culture & Induction: Grow MRSA strain (e.g., COL, N315) to mid-log phase (OD600 ~0.5). Divide culture. Treat one aliquot with inducing β-lactam (e.g., 0.5 µg/ml oxacillin for MecR1, 0.1 µg/ml penicillin G for BlaR1). Maintain an uninduced control.
- Sampling: Collect 1 ml aliquots at T=0, 5, 15, 30, 60, 120 minutes post-induction. Pellet cells rapidly (13,000 rpm, 1 min).
- Lysis: Resuspend pellets in 100 µl SDS-PAGE loading buffer with 1mM PMSF. Heat at 95°C for 10 minutes. Use lysostaphin (200 µg/ml) pre-treatment for efficient lysis if needed.
- Detection: Perform SDS-PAGE (12% gel), transfer to PVDF membrane. Probe with:
- Primary antibodies: Anti-BlaI, Anti-MecI, Anti-BlaR1 (C-terminal epitope), Anti-MecR1 (C-terminal epitope).
- Secondary antibody: HRP-conjugated anti-rabbit IgG.
- Analysis: Quantify band intensity using chemiluminescence imaging to track repressor disappearance and sensor fragmentation.
Protocol 2: In Vivo Signal Duration and Recovery Assay using Reporter Gene
- Objective: To quantify the kinetics of induction and signal recovery for each system.
- Method:
- Reporter Strain Construction: Fuse promoter of blaZ or mecA to a stable GFP gene in a replicating plasmid. Transform into relevant S. aureus strain.
- Induction & Wash-Out: Induce culture as in Protocol 1. After 60 minutes, harvest cells by centrifugation, wash 3x with warm, antibiotic-free medium.
- Flow Cytometry: Resuspend washed cells in fresh medium and continue incubation. Measure GFP fluorescence (FITC channel) via flow cytometry at 30-minute intervals for 6-8 hours.
- Data Modeling: Plot mean fluorescence intensity vs. time. Fit curves to determine induction rate, peak time, and recovery half-time (time for signal to decay to 50% of peak).
Protocol 3: Cycloheximide Chase to Determine Sensor Protein Half-Life
- Objective: To directly compare the post-induction stability of BlaR1 vs. MecR1.
- Method:
- Induction: Induce cultures as in Protocol 1.
- Translation Inhibition: At peak induction (T=30 min), add cycloheximide (100 µg/ml) to inhibit further protein synthesis.
- Sampling: Collect samples immediately before (T=30) and after (T=45, 60, 90, 120 min) cycloheximide addition.
- Analysis: Process samples via immunoblotting (Protocol 1) for BlaR1/MecR1. Use a constitutively expressed protein (e.g., GyrA) as loading control. Plot residual full-length sensor signal to estimate half-life.
Pathway and Conceptual Diagrams
Title: Comparative signaling pathways of BlaR1 and MecR1 in MRSA
Title: Workflow for analyzing sensor degradation and signal recovery kinetics
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagent Solutions for BlaR1/MecR1 Signal Transduction Research
| Reagent / Material | Function / Purpose | Example & Notes |
|---|---|---|
| Inducing Antibiotics | To specifically activate BlaR1 or MecR1 pathways. | Benzylpenicillin (Pen G): For BlaR1. Oxacillin/Cefoxitin: For MecR1. Use at sub-MIC concentrations (0.1-1 µg/ml). |
| Protease Inhibitors | To stabilize protein fragments during lysis and study proteolytic events. | PMSF (1 mM), Protease Inhibitor Cocktail (EDTA-free): Added to lysis buffer to halt degradation ex vivo. |
| Epitope-Specific Antibodies | To detect full-length sensors, cleavage fragments, and repressors via immunoblotting. | Anti-BlaR1 (C-term), Anti-MecR1 (C-term), Anti-BlaI, Anti-MecI. Crucial for distinguishing cleavage products. |
| Lysozyme/Lysostaphin | For efficient lysis of S. aureus cell walls to extract proteins. | Lysostaphin (200 µg/ml): Highly specific for S. aureus peptidoglycan. Pre-treat cells before SDS buffer. |
| Cycloheximide | Eukaryotic translation inhibitor used in chase experiments in S. aureus. | Used at high concentrations (100-200 µg/ml) to block new protein synthesis for half-life studies. |
| Reporter Plasmid | To quantify promoter activity and signal dynamics in live cells. | pCN51-P_{blaZ/mecA}-GFP: Shuttle plasmid with constitutive antibiotic resistance and inducible GFP. |
| Clp Protease Inhibitors/Modulators | To investigate the role of specific degradation machinery. | ADEP analogs (activators of ClpP): Experimental tools to probe Clp-dependent degradation of repressors/sensors. |
| Chromogenic β-lactam Substrate | To measure β-lactamase (BlaZ) activity as a functional output. | Nitrocefin: Changes color from yellow to red upon hydrolysis. Used in kinetic assays. |
This whitepaper situates the analysis of tetracycline (TetR) and macrolide (MphR) repressor systems within a broader thesis framework investigating BlaR1 fragmentation and turnover as a novel paradigm for antimicrobial resistance (AMR) signal recovery. Unlike β-lactam-sensing BlaR1, a membrane-bound sensor-transducer that undergoes irreversible proteolytic cleavage to derepress resistance genes, TetR and MphR represent cytoplasmic, ligand-binding transcriptional regulators. A comparative analysis highlights fundamental mechanistic dichotomies in signal perception, propagation, and resetting, informing strategies to interfere with resistance pathways.
Core System Mechanisms & Quantitative Data
| Feature | BlaR1 (β-lactam) | TetR (Tetracycline) | MphR(A) (Macrolide) |
|---|---|---|---|
| Regulator Type | Membrane-bound sensor-transducer (MecR1 homolog) | Cytoplasmic, homodimeric helix-turn-helix repressor | Cytoplasmic, homodimeric repressor (MarR family) |
| Inducing Ligand | β-Lactam antibiotics (covalent acylation) | Tetracycline-Mg²⁺ complex (non-covalent) | Erythromycin, other 14- and 15-membered macrolides (non-covalent) |
| DNA Binding | BlaI repressor (cleaved/inactivated by BlaR1) | Direct binding to tet operator (O) sequences | Direct binding to inverted repeats in mphR promoter region |
| Signal Transmission | Cysteine protease domain activation → BlaI cleavage | Allosteric conformational change upon ligand binding | Allosteric conformational change upon ligand binding |
| System Reset | Irreversible; requires new synthesis (proteolytic turnover). Core to fragmentation thesis. | Reversible; ligand dissociation restores DNA affinity. | Reversible; ligand dissociation restores DNA affinity. |
| Key Derepressed Gene | blaZ (β-lactamase) | tetA (efflux pump) | mphA (macrolide phosphotransferase) |
| Dissociation Constant (Kd) | N/A (covalent) | ~1 nM for Tc-Mg²⁺ to operator DNA (repressed state) | ~10-100 nM for Erythromycin (induction EC₅₀) |
| Operon Structure | blaR1-blaI-blaZ | tetR-tetA (or variants) | mphR-mphA |
Experimental Protocols for Key Analyses
Protocol: Electrophoretic Mobility Shift Assay (EMSA) for TetR/MphR DNA Binding
Objective: Quantify repressor-operator affinity and ligand-induced dissociation.
- Protein Purification: Express His₆-tagged TetR/MphR in E. coli and purify via Ni²⁺-NTA chromatography.
- DNA Probe Preparation: PCR amplify or anneal fluorescently labeled (e.g., FAM, Cy5) oligonucleotides containing the cognate operator sequence (tetO or mphO).
- Binding Reaction: Incubate purified protein (0-500 nM range) with fixed concentration of labeled DNA probe (1-10 nM) in binding buffer (20 mM Tris-HCl pH 8.0, 50 mM KCl, 5% glycerol, 1 mM DTT, 100 μg/mL BSA) ± inducer (Tc-Mg²⁺ or erythromycin at 10-100 μM). Include non-specific competitor DNA (e.g., poly(dI-dC)).
- Electrophoresis: Resolve complexes on a pre-run 6-8% native polyacrylamide gel in 0.5x TBE at 4°C, 100V for 60-90 min.
- Analysis: Visualize using a fluorescence gel scanner. Quantify free vs. bound DNA to determine apparent Kd and ligand IC₅₀.
Protocol: Isothermal Titration Calorimetry (ITC) for Ligand Binding
Objective: Directly measure thermodynamic parameters of inducer-repressor interaction.
- Sample Preparation: Dialyze purified repressor (TetR/MphR) and ligand (Tc-Mg²⁺ or erythromycin) into identical degassed buffer (e.g., 20 mM HEPES pH 7.5, 150 mM NaCl).
- Instrument Setup: Load the repressor solution (50-100 μM) into the sample cell. Fill the syringe with ligand solution (10x concentrated relative to protein).
- Titration: Perform a series of automated injections (e.g., 19 x 2 μL) of ligand into the protein solution while continuously measuring the heat change required to maintain temperature equilibrium.
- Data Fitting: Integrate heat peaks, subtract control titrations (ligand into buffer), and fit the binding isotherm to a single-site model to derive Kd, ΔH (enthalpy), ΔS (entropy), and stoichiometry (n).
Protocol: In Vivo Reporter Assay for Induction Kinetics
Objective: Monitor real-time derepression dynamics and contrast reversibility with Bla systems.
- Reporter Construction: Clone the promoter-operator region of tetA or mphA upstream of a promoterless lacZ or gfp gene on a reporter plasmid.
- Strain Generation: Transform reporter plasmid into a host strain harboring the native or plasmid-encoded tetR/mphR gene.
- Induction & Measurement: Grow cultures to mid-log phase. Add inducer (Tc or Ery) and monitor β-galactosidase activity (Miller assay) or fluorescence (GFP) over time (0-180 min). For reversibility test, wash cells post-induction to remove antibiotic and continue monitoring reporter output.
- Data Analysis: Plot induction kinetics. Compare with analogous Bla system assays where induction is irreversible.
Visualization of Signaling Pathways
Title: Reversible Induction in TetR and MphR Systems
Title: BlaR1 Irreversible Proteolysis and Turnover
The Scientist's Toolkit: Research Reagent Solutions
| Reagent/Material | Function & Application in TetR/MphR Research |
|---|---|
| Purified TetR/MphR Protein (His-tagged) | Essential for in vitro studies (EMSA, ITC, crystallography) to characterize DNA/ligand binding without cellular complexity. |
| Fluorescently-labeled tetO/mphO DNA Oligos | High-sensitivity probes for EMSA and fluorescence anisotropy assays to quantify repressor-operator affinity and kinetics. |
| Isothermal Titration Calorimeter (ITC) | Instrument for direct, label-free measurement of binding thermodynamics between repressor and antibiotic inducers. |
| Reporter Plasmid (PtetA/mphA-GFP/lacZ) | For in vivo monitoring of induction kinetics, promoter strength, and system reversibility in live bacterial cells. |
| Surface Plasmon Resonance (SPR) Chip with immobilized DNA | Enables real-time kinetic analysis (association/dissociation rates) of repressor binding to operator sequences ± ligands. |
| Anhydrotetracycline (aTc) | A potent, stable tetracycline analog often used as a high-affinity inducer in experimental systems to minimize antibiotic side effects. |
| Size Exclusion Chromatography (SEC) Column | To assess ligand-induced conformational changes via shifts in oligomeric state or Stokes radius of the repressor protein. |
| Crystallization Screens (e.g., Hampton Research) | For obtaining high-resolution structures of apo- and ligand-bound repressor complexes to elucidate allosteric mechanisms. |
Within the broader thesis on BlaR1 fragmentation and signal recovery, understanding the distinction between targeting BlaR1 turnover and inhibiting β-lactamase is critical. BlaR1 is the transmembrane sensor-transducer responsible for initiating the bacterial resistance response to β-lactam antibiotics. Inhibiting the β-lactamase enzyme itself, which hydrolyzes β-lactams, is a well-established but increasingly challenged strategy due to enzyme evolution. In contrast, targeting the turnoff or signal recovery mechanism of BlaR1—specifically, its fragmentation and removal from the membrane—presents a novel therapeutic avenue aimed at permanently locking the sensor in an "on" state, leading to constitutive, detrimental expression of resistance genes and potentially restoring antibiotic efficacy.
Core Mechanisms: BlaR1 Signaling vs. β-Lactamase Action
The key difference lies in the target's function: β-lactamase is a defensive enzyme, while BlaR1 is a signaling protein controlling gene expression.
BlaR1 Signaling Pathway: Upon binding a β-lactam antibiotic, the sensor domain of BlaR1 (a penicillin-binding protein homolog) covalently acylates. This event triggers an intramembrane proteolysis event, liberating a cytosolic fragment that functions as a zinc-dependent transcription factor. This fragment binds to the promoter region of the bla operon, inducing transcription of β-lactamase (BlaZ) and, in some systems, BlaR1 itself. Signal termination ("turnoff") involves further proteolytic processing (e.g., ClpXP-mediated degradation) of the cytosolic fragment, allowing the system to reset.
β-Lactamase Function: The BlaZ enzyme secreted into the periplasm hydrolytically cleaves the β-lactam ring of incoming antibiotics, rendering them inactive before they can inhibit essential penicillin-binding proteins (PBPs) involved in cell wall synthesis.
Diagram: BlaR1 Signaling vs. β-Lactamase Inhibition Pathways
Diagram Title: BlaR1 Signal Transduction and β-Lactamase Action
Quantitative Comparison of Therapeutic Targets
The table below summarizes the fundamental differences between the two strategies.
Table 1: Target Comparison - BlaR1 Turnoff vs. β-Lactamase
| Parameter | Targeting β-Lactamase (BlaZ) | Targeting BlaR1 Turnoff/Signal Recovery |
|---|---|---|
| Primary Objective | Neutralize the hydrolytic enzyme to protect the antibiotic. | Disrupt signal termination to cause constitutive, futile resistance gene expression. |
| Target Molecule | Secreted/periplasmic enzyme. | Transmembrane sensor/transducer & cytosolic proteases. |
| Molecular Mechanism | Competitive or irreversible inhibition of the active site. | Inhibition of proteolytic fragmentation (e.g., ClpXP) or stabilization of the active cytosolic fragment. |
| Therapeutic Outcome | Restores activity of a specific β-lactam antibiotic. | Potentially restores activity of all β-lactams sensed by BlaR1. |
| Resistance Risk | High. Mutations in β-lactamase can evade inhibitors. | Theoretically Lower. Targeting host protease machinery may be less mutable. |
| Stage of Intervention | Downstream. Acts after resistance gene expression. | Upstream. Prevents or dysregulates the induction signal. |
| Experimental Readout | MIC (Minimum Inhibitory Concentration) reduction in combo assays. | β-lactamase promoter activity (e.g., reporter assays), fragment persistence (Western), bacterial lysis kinetics. |
Experimental Protocols for BlaR1 Turnoff Research
Protocol: Monitoring BlaR1 Cytosolic Fragment Turnover
Objective: To assess the stability and degradation kinetics of the BlaR1-derived cytosolic transcription factor fragment.
Materials: See "Scientist's Toolkit" below. Procedure:
- Culture & Induction: Grow a laboratory strain of Staphylococcus aureus (e.g., RN4220 harboring a native or plasmid-borne bla operon) to mid-log phase (OD600 ~0.5).
- Pulse-Chase: Induce the system with a sub-MIC concentration of a potent inducer (e.g., 0.1 µg/ml cefoxitin) for 10 minutes (pulse).
- Chase: Rapidly remove the inducer by washing cells 3x in warm, antibiotic-free medium. Resuspend in fresh medium. Aliquot samples at time points: 0, 5, 15, 30, 60 minutes post-wash.
- Lysis & Fractionation: Pellet samples. Lyse cells using a bead-beater or lysostaphin in the presence of protease inhibitors (omit for degradation studies). Remove cell debris by centrifugation. Optionally, perform membrane fractionation via ultracentrifugation (100,000 x g, 1h) to isolate the cytosolic fraction.
- Immunoblotting: Separate proteins by SDS-PAGE (12% gel). Transfer to PVDF membrane. Probe with a custom polyclonal antibody specific for the N-terminal or cytosolic domain of BlaR1 (anti-BlaR1cyt).
- Quantification: Use chemiluminescence and densitometry to quantify the intensity of the cytosolic fragment band over time. Plot fragment persistence (%) vs. time. Fit the curve to determine half-life (t1/2).
Protocol: Reporter Assay for Constitutive Signal Activation
Objective: To screen for compounds that inhibit BlaR1 fragment turnover by measuring sustained promoter activity.
Procedure:
- Reporter Strain Construction: Fuse the promoter of the β-lactamase gene (blaP) or the bla operon to a reporter gene (e.g., lacZ, gfp, luxABCDE) in an appropriate Gram-positive vector. Transform into S. aureus.
- Compound Screening: In a 96-well plate, grow the reporter strain with sub-inhibitory concentrations of test compounds (potential ClpXP inhibitors or fragment stabilizers) for 1 hour.
- Signal Induction & Monitoring: Add a brief, low-concentration pulse of a β-lactam inducer (e.g., 0.05 µg/ml methicillin) for 20 minutes. Wash cells to remove the inducer.
- Readout: Continue incubation with the test compound but without the inducer. Measure reporter signal (e.g., fluorescence for GFP, luminescence for luciferase) every 30 minutes for 4-6 hours.
- Data Analysis: Compare signal decay kinetics between treated and untreated wells. A successful "turnoff inhibitor" will result in sustained high signal post-induction pulse, indicating failure to reset the system.
Diagram: Experimental Workflow for Fragment Turnover Analysis
Diagram Title: BlaR1 Fragment Turnover Assay Workflow
The Scientist's Toolkit: Key Research Reagent Solutions
Table 2: Essential Materials for BlaR1 Turnoff Research
| Reagent/Material | Function/Biological Role | Example or Specification |
|---|---|---|
| Inducer β-Lactams | Acylate the sensor domain of BlaR1, initiating the signaling cascade. | Cefoxitin, Methicillin, Cloxacillin (low, sub-MIC concentrations). |
| Anti-BlaR1 Cytosolic Domain Antibody | Detects the liberated transcription factor fragment and full-length BlaR1 in immunoblots. | Custom polyclonal antibody raised against a purified N-terminal fragment. |
| Protease Inhibitor Cocktail | Preserves protein fragments during cell lysis for snapshot analyses. | EDTA-free cocktail (to preserve zinc-dependent proteases) for turnover studies. |
| ClpXP Inhibitor (Positive Control) | Pharmacologically inhibits the protease responsible for fragment degradation, validating the assay. | ADEP (Acyldepsipeptide) analogs (use with caution due to pleiotropic effects). |
| Reporter Plasmid (Pbla-reporter) | Quantifies the transcriptional output of the BlaR1 signaling pathway in real-time. | S. aureus shuttle vector with blaP promoter driving gfp or luc. |
| Zinc Chelator | Investigates the role of the zinc-binding domain in fragment stability and function. | 1,10-Phenanthroline (cell-permeable). |
| Membrane Fractionation Kit | Isolates membrane-bound BlaR1 from the cytosolic fragment for localization studies. | Commercial kits for Gram-positive bacteria. |
| BlaR1 Knockout/ Mutant Strain | Provides a negative control for signaling and fragment detection. | S. aureus strain with in-frame deletion of blaR1. |
Targeting BlaR1 turnoff represents a paradigm shift from inhibiting the resistance factor (β-lactamase) to manipulating the sensory apparatus that controls its production. This strategy, framed within signal recovery research, aims to exhaust the bacterial cell via futile overproduction of resistance proteins, potentially leading to restored antibiotic susceptibility and slower resistance development. Future work requires high-throughput screening for specific BlaR1 fragment stabilizers or ClpXP inhibitors, combined with structural studies of the BlaR1 fragment-protease complex, to translate this novel concept into viable therapeutic leads.
Conclusion
The fragmentation and proteasomal turnover of BlaR1 is not merely a cleanup mechanism but a fundamental, regulated reset switch essential for the adaptive response of bacteria to β-lactams. This detailed mechanistic understanding, validated against related systems, underscores a critical vulnerability in the resistance pathway. Unlike targeting the downstream β-lactamase, which faces an enzyme multiplicity problem, disrupting BlaR1 inactivation offers a novel strategy to 'freeze' the resistance signal, preventing phenotypic recovery and potentially re-sensitizing bacteria. Future research must focus on high-resolution structural studies of BlaR1 degradation intermediates and the development of specific small-molecule inhibitors that block its proteasomal recognition, paving the way for a new class of antibiotic adjuvants that target signal transduction over enzymatic activity.